

Neocortical long-term potentiation and long-term depression: site of expression investigated by infrared-guided laser stimulation
- PMID: 12196579
- PMCID: PMC6758004
- DOI: 10.1523/JNEUROSCI.22-17-07558.2002
Neocortical long-term potentiation and long-term depression: site of expression investigated by infrared-guided laser stimulation
Abstract
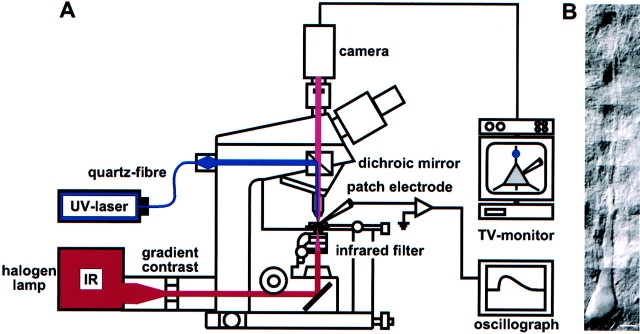
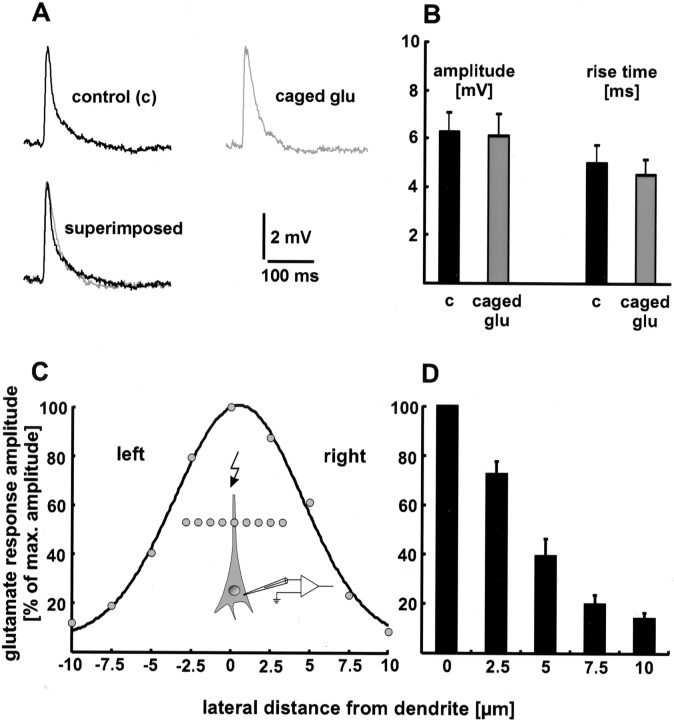
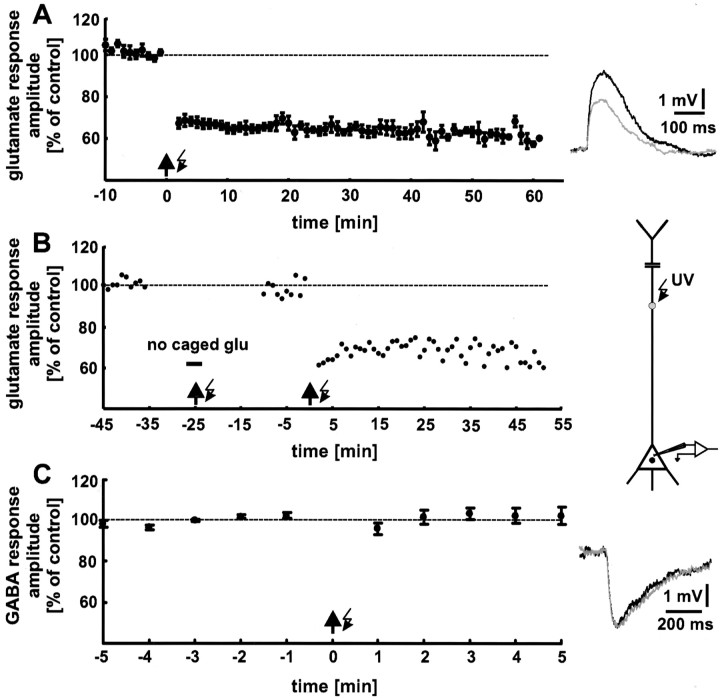
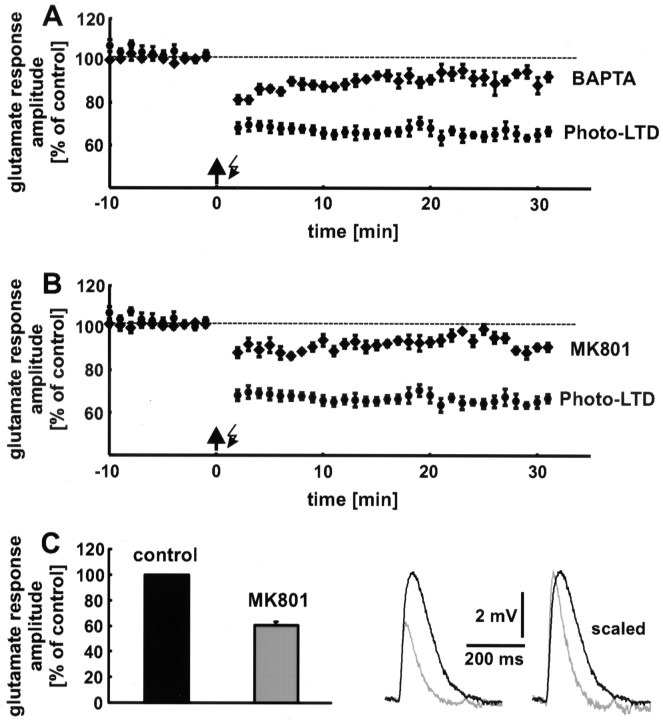
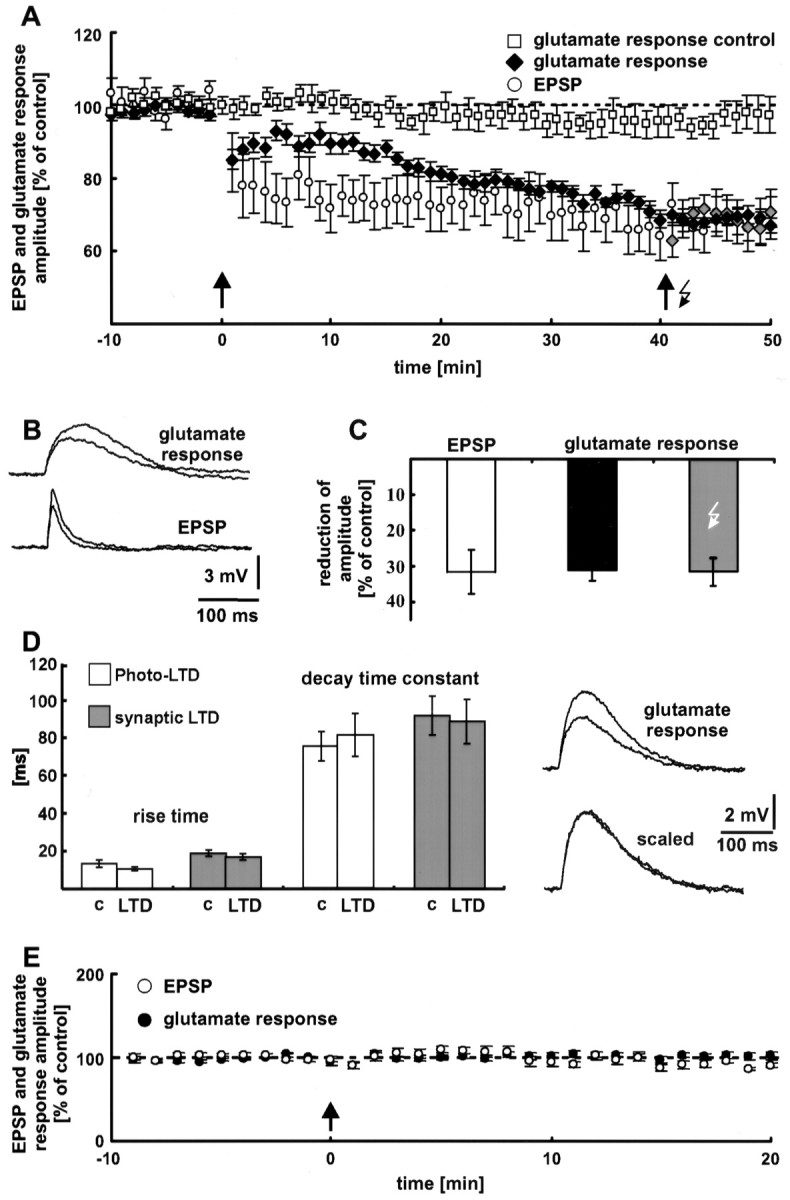
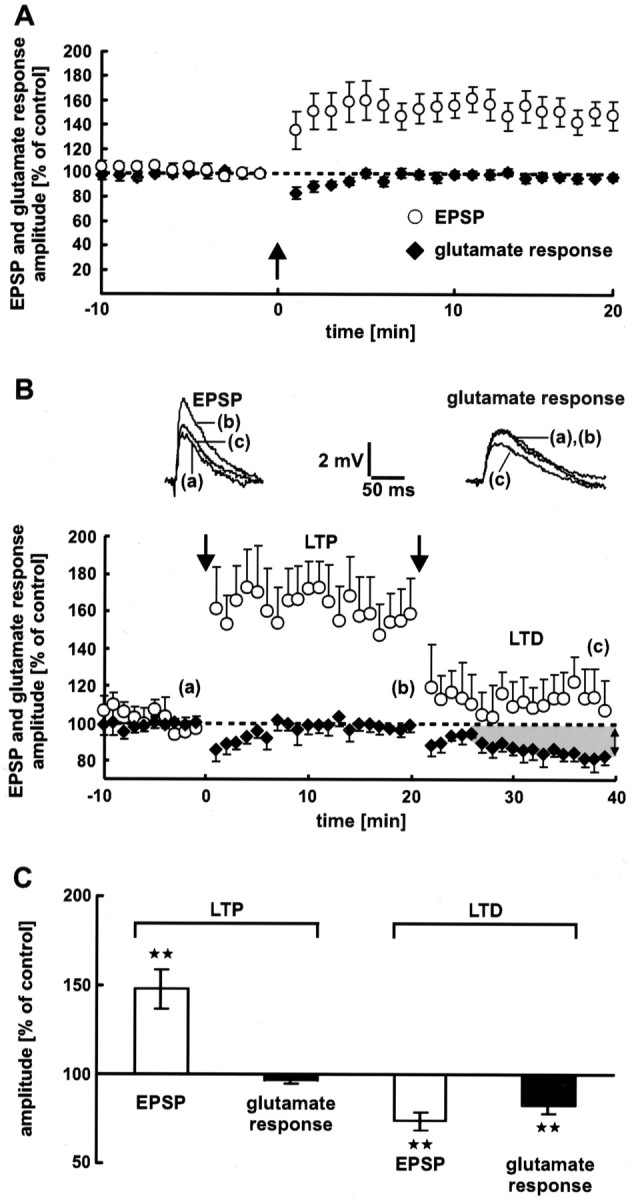
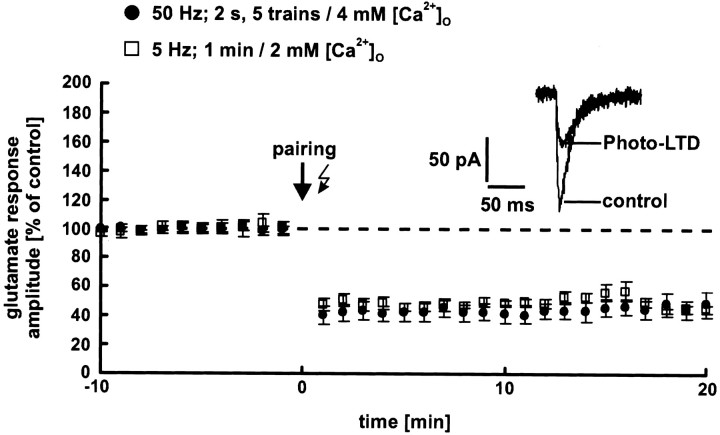
The synaptic site of expression of long-term potentiation (LTP) and long-term depression (LTD) is still a matter of debate. To address the question of presynaptic versus postsynaptic expression of neocortical LTP and LTD in a direct approach, we measured the glutamate sensitivity of apical dendrites of layer 5 pyramidal neurons during LTP and LTD. We used infrared-guided laser stimulation to release glutamate from its "caged" form with high spatial and temporal resolution. Responses to photolytically released glutamate and synaptically evoked EPSPs were recorded with patch-clamp pipettes from the neuronal somata. LTP and LTD could be induced by electrical stimulation at the same synapses in succession. The NMDA receptor-dependent LTD was accompanied by a decrease in the dendritic glutamate sensitivity, suggesting a postsynaptic expression of neocortical LTD. In contrast, LTP was never accompanied by a change in the dendritic glutamate sensitivity. A possible explanation for this finding is a presynaptic expression of neocortical LTP. Another set of experiments corroborated these results: Photolytic application of glutamate with a frequency of 5 Hz caused a long-lasting Ca2+ and NMDA receptor-dependent decrease in the dendritic glutamate sensitivity. In contrast, LTP of dendritic glutamate sensitivity was never induced by photostimulation, despite several experimental modifications to prevent washout of the induction mechanism and to induce a stronger postsynaptic Ca2+ influx. In conclusion, our findings provide strong evidence for a postsynaptic expression of neocortical LTD and favor a primarily presynaptic locus of neocortical LTP.
Figures
Similar articles
-
Precisely localized LTD in the neocortex revealed by infrared-guided laser stimulation.Science. 1999 Oct 1;286(5437):110-3. doi: 10.1126/science.286.5437.110. Science. 1999. PMID: 10506556
-
Coincident spiking activity induces long-term changes in inhibition of neocortical pyramidal cells.J Neurosci. 2001 Oct 15;21(20):8270-7. doi: 10.1523/JNEUROSCI.21-20-08270.2001. J Neurosci. 2001. PMID: 11588198 Free PMC article.
-
Two forms of synaptic plasticity with distinct dependence on age, experience, and NMDA receptor subtype in rat visual cortex.J Neurosci. 2003 Jul 23;23(16):6557-66. doi: 10.1523/JNEUROSCI.23-16-06557.2003. J Neurosci. 2003. PMID: 12878697 Free PMC article.
-
The role of dendritic filtering in associative long-term synaptic plasticity.Learn Mem. 1999 Sep-Oct;6(5):422-47. doi: 10.1101/lm.6.5.422. Learn Mem. 1999. PMID: 10541464 Review.
-
Theta-burst LTP.Brain Res. 2015 Sep 24;1621:38-50. doi: 10.1016/j.brainres.2014.10.034. Epub 2014 Oct 27. Brain Res. 2015. PMID: 25452022 Free PMC article. Review.
Cited by
-
Optogenetic evocation of field inhibitory postsynaptic potentials in hippocampal slices: a simple and reliable approach for studying pharmacological effects on GABAA and GABAB receptor-mediated neurotransmission.Front Cell Neurosci. 2014 Jan 22;8:2. doi: 10.3389/fncel.2014.00002. eCollection 2014. Front Cell Neurosci. 2014. PMID: 24478627 Free PMC article.
-
A calcium-based plasticity model for predicting long-term potentiation and depression in the neocortex.Nat Commun. 2022 Jun 1;13(1):3038. doi: 10.1038/s41467-022-30214-w. Nat Commun. 2022. PMID: 35650191 Free PMC article.
-
Fully-primed slowly-recovering vesicles mediate presynaptic LTP at neocortical neurons.Proc Natl Acad Sci U S A. 2023 Oct 24;120(43):e2305460120. doi: 10.1073/pnas.2305460120. Epub 2023 Oct 19. Proc Natl Acad Sci U S A. 2023. PMID: 37856547 Free PMC article.
-
Synaptic mechanisms for plasticity in neocortex.Annu Rev Neurosci. 2009;32:33-55. doi: 10.1146/annurev.neuro.051508.135516. Annu Rev Neurosci. 2009. PMID: 19400721 Free PMC article. Review.
-
Early excitatory-inhibitory cortical modifications following skill learning are associated with motor memory consolidation and plasticity overnight.Nat Commun. 2024 Jan 30;15(1):906. doi: 10.1038/s41467-024-44979-9. Nat Commun. 2024. PMID: 38291029 Free PMC article.
References
-
- Andreasen M, Hablitz JJ. Paired-pulse facilitation in the dentate gyrus: a patch-clamp study in rat hippocampus in vitro. J Neurophysiol. 1994;72:326–336. - PubMed
-
- Arancio O, Kandel ER, Hawkins RD. Activity-dependent long-term enhancement of transmitter release by presynaptic 3′,5′-cyclic GMP in cultured hippocampal neurons. Nature. 1995;376:74–80. - PubMed
-
- Arancio O, Kiebler M, Lee CJ, Lev-Ram V, Tsien RY, Kandel ER, Hawkins RD. Nitric oxide acts directly in the presynaptic neuron to produce long-term potentiation in cultured hippocampal neurons. Cell. 1996;87:1025–1035. - PubMed
-
- Barria A, Muller D, Derkach V, Griffith LC, Soderling TR. Regulatory phosphorylation of AMPA-type glutamate receptors by CaM-KII during long-term potentiation. Science. 1997;276:2042–2045. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous