

Distinct actions of Emx1, Emx2, and Pax6 in regulating the specification of areas in the developing neocortex
- PMID: 12196586
- PMCID: PMC6757966
- DOI: 10.1523/JNEUROSCI.22-17-07627.2002
Distinct actions of Emx1, Emx2, and Pax6 in regulating the specification of areas in the developing neocortex
Abstract
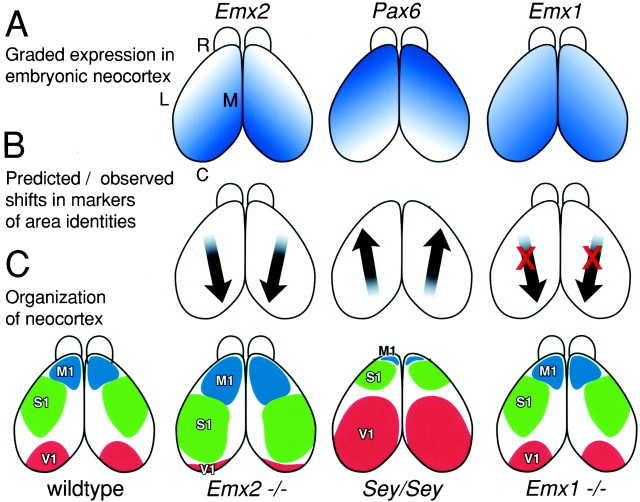
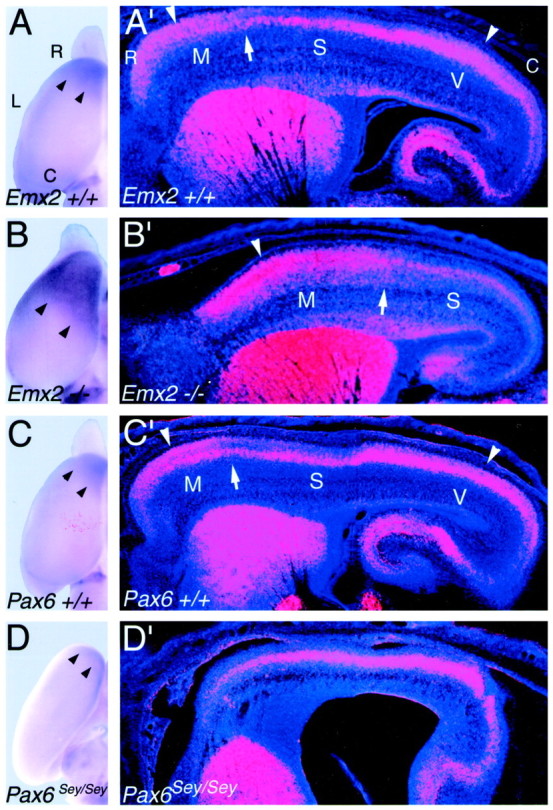
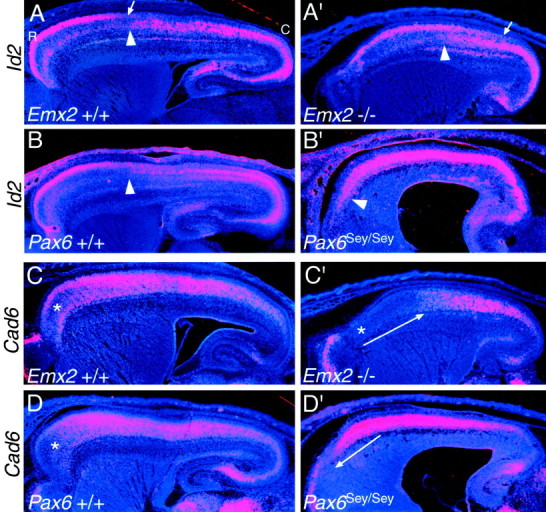
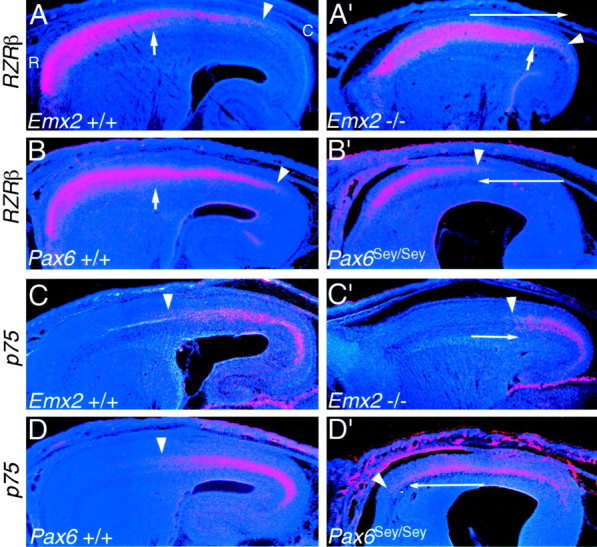
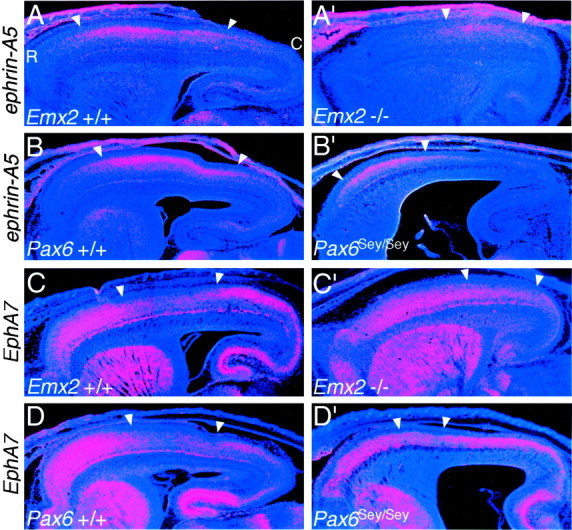
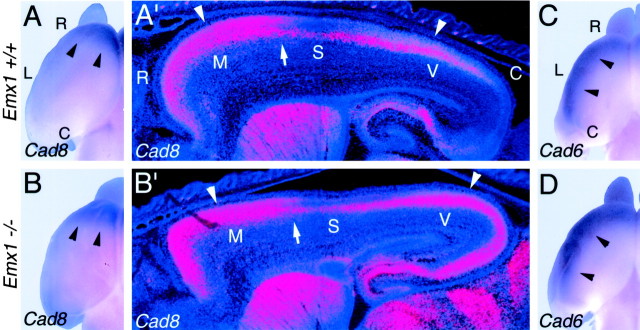
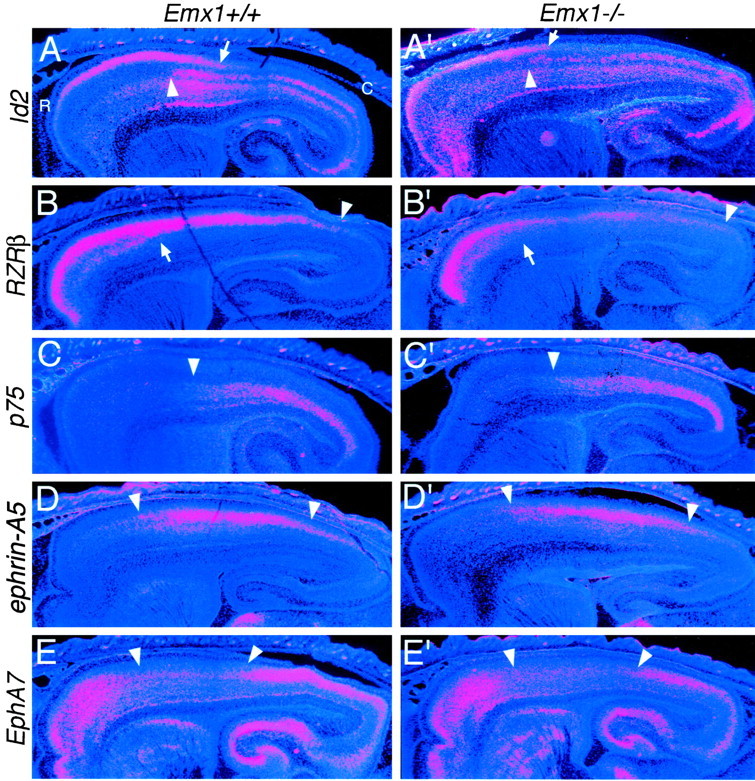
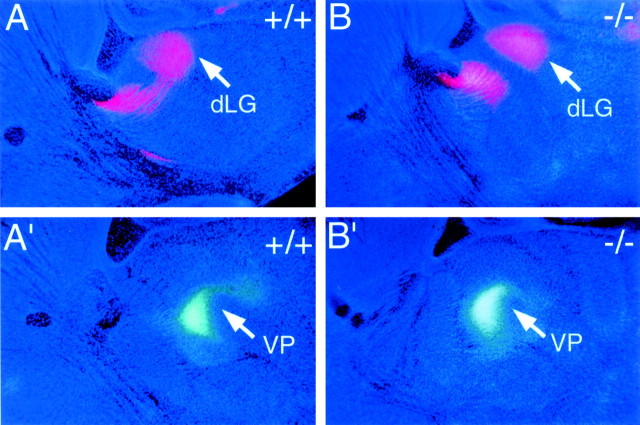
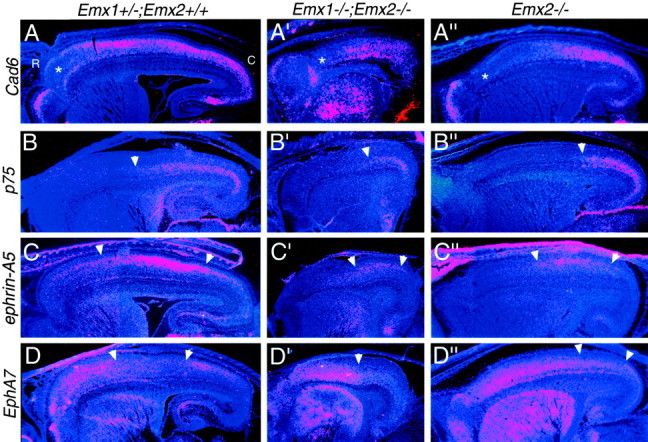
The mammalian neocortex is organized into subdivisions referred to as areas that are distinguished from one another by differences in architecture, axonal connections, and function. The transcription factors EMX1, EMX2, and PAX6 have been proposed to regulate arealization. Emx1 and Emx2 are expressed by progenitor cells in a low rostrolateral to high caudomedial gradient across the embryonic neocortex, and Pax6 is expressed in a high rostrolateral to low caudomedial gradient. Recent evidence has suggested that EMX2 and PAX6 have a role in the genetic regulation of arealization. Here we use a panel of seven genes (Cad6, Cad8, Id2, RZRbeta, p75, EphA7, and ephrin-A5) representative of a broad range of proteins as complementary markers of positional identity to obtain a more thorough assessment of the suggested roles for EMX2 and PAX6 in arealization, and in addition to assess the proposed but untested role for EMX1 in arealization. Orderly changes in the size and positioning of domains of marker expression in Emx2 and Pax6 mutants strongly imply that rostrolateral areas (motor and somatosensory) are expanded, whereas caudomedial areas (visual) are reduced in Emx2 mutants and that opposite effects occur in Pax6 mutants, consistent with their opposing gradients of expression. In contrast, patterns of marker expression, as well as the distribution of area-specific thalamocortical projections, appear normal in Emx1 mutants, indicating that they do not exhibit changes in arealization. This lack of a defined role for EMX1 in arealization is supported by our finding of similar shifts in patterns of marker expression in Emx1; Emx2 double mutants as in Emx2 mutants. Thus, our findings indicate that EMX2 and PAX6 regulate, in opposing manners, arealization of the neocortex and impart positional identity to cortical cells, whereas EMX1 appears not to have a role in this process.
Figures
References
-
- Bishop KM, Goudreau G, O'Leary DDM. Regulation of area identity in the mammalian neocortex by Emx2 and Pax6. Science. 2000;288:344–349. - PubMed
-
- Caric D, Gooday D, Hill RE, McConnell SK, Price DJ. Determination of the migratory capacity of embryonic cortical cells lacking the transcription factor Pax-6. Development. 1997;124:5087–5096. - PubMed
-
- Cecchi C, Boncinelli E. Emx homeogenes and mouse brain development. Trends Neurosci. 2000;23:347–352. - PubMed
-
- Chenn A, Braisted JE, McConnell SK, O'Leary DDM. Development of the cerebral cortex: mechanisms controlling cell fate, laminar and areal patterning, and axonal connectivity. In: Dowan WM, Zipursky L, Jessell T, editors. Molecular and cellular approaches to neural development. Oxford UP; New York: 1997. pp. 440–473.
-
- Cohen-Tannoudji M, Babinet C, Wassef M. Early determination of a mouse somatosensory cortex marker. Nature. 1994;368:460–463. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous