

The antiangiogenic activity of cleaved high molecular weight kininogen is mediated through binding to endothelial cell tropomyosin
- PMID: 12196635
- PMCID: PMC129426
- DOI: 10.1073/pnas.192668299
The antiangiogenic activity of cleaved high molecular weight kininogen is mediated through binding to endothelial cell tropomyosin
Abstract
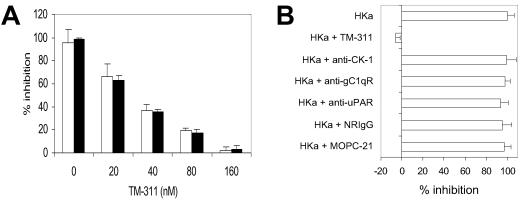
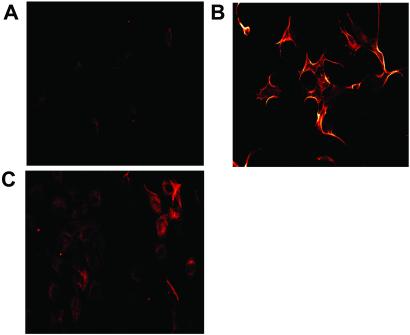
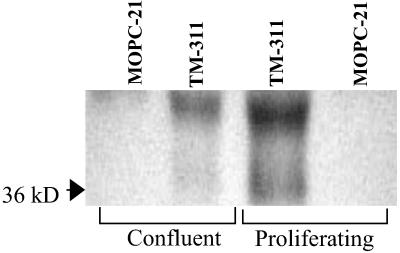
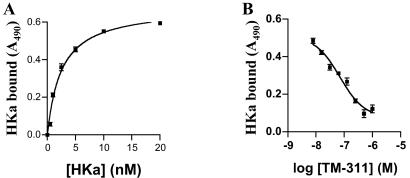
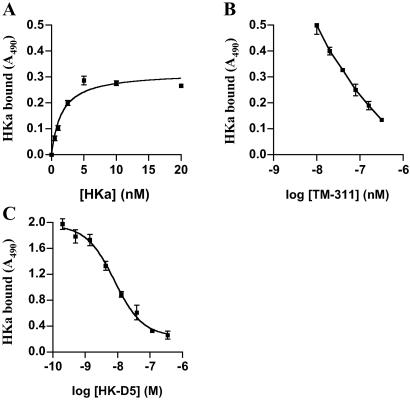
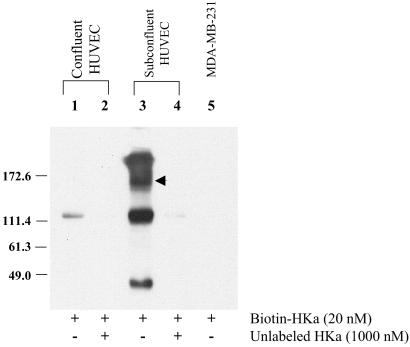
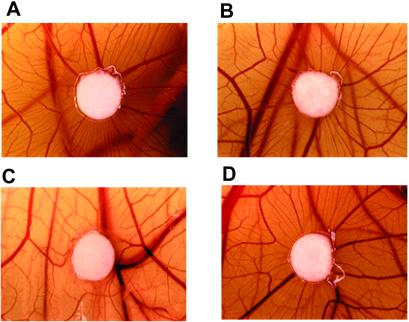
Conformationally altered proteins and protein fragments derived from the extracellular matrix and hemostatic system may function as naturally occurring angiogenesis inhibitors. One example of such a protein is cleaved high molecular weight kininogen (HKa). HKa inhibits angiogenesis by inducing apoptosis of proliferating endothelial cells, effects mediated largely by HKa domain 5. However, the mechanisms underlying the antiangiogenic activity of HKa have not been characterized, and its binding site on proliferating endothelial cells has not been defined. Here, we report that the induction of endothelial cell apoptosis by HKa, as well as the antiangiogenic activity of HKa in the chick chorioallantoic membrane, was inhibited completely by antitropomyosin monoclonal antibody TM-311. TM-311 also blocked the high-affinity Zn2+-dependent binding of HKa to both purified tropomyosin and proliferating endothelial cells. Confocal microscopic analysis of endothelial cells stained with monoclonal antibody TM-311, as well as biotin labeling of cell surface proteins on intact endothelial cells, revealed that tropomyosin exposure was enhanced on the surface of proliferating cells. These studies demonstrate that the antiangiogenic effects of HKa depend on high-affinity binding to endothelial cell tropomyosin.
Figures
Similar articles
-
Inhibition of angiogenesis by two-chain high molecular weight kininogen (HKa) and kininogen-derived polypeptides.Can J Physiol Pharmacol. 2002 Feb;80(2):85-90. doi: 10.1139/y02-011. Can J Physiol Pharmacol. 2002. PMID: 11934260
-
Inhibition of angiogenesis by antibody blocking the action of proangiogenic high-molecular-weight kininogen.J Thromb Haemost. 2003 Jan;1(1):164-70. doi: 10.1046/j.1538-7836.2003.00025.x. J Thromb Haemost. 2003. PMID: 12871554
-
Kininostatin as an antiangiogenic inhibitor: what we know and what we do not know.Int Immunopharmacol. 2002 Dec;2(13-14):1931-40. doi: 10.1016/s1567-5769(02)00172-8. Int Immunopharmacol. 2002. PMID: 12489806 Review.
-
Two-chain high molecular weight kininogen induces endothelial cell apoptosis and inhibits angiogenesis: partial activity within domain 5.FASEB J. 2000 Dec;14(15):2589-600. doi: 10.1096/fj.99-1025com. FASEB J. 2000. PMID: 11099478
-
Inhibition of angiogenesis by cleaved high molecular weight kininogen (HKa) and HKa domain 5.Curr Cancer Drug Targets. 2005 Nov;5(7):519-28. doi: 10.2174/156800905774574039. Curr Cancer Drug Targets. 2005. PMID: 16305348 Review.
Cited by
-
Angiogenesis inhibitors found within the haemostasis pathway.J Cell Mol Med. 2005 Apr-Jun;9(2):286-302. doi: 10.1111/j.1582-4934.2005.tb00356.x. J Cell Mol Med. 2005. PMID: 15963250 Free PMC article. Review.
-
Cell death pathways and viruses: Role of microRNAs.Mol Ther Nucleic Acids. 2021 Mar 19;24:487-511. doi: 10.1016/j.omtn.2021.03.011. eCollection 2021 Jun 4. Mol Ther Nucleic Acids. 2021. PMID: 33898103 Free PMC article. Review.
-
The inhibitory effect of HKa in endothelial cell tube formation is mediated by disrupting the uPA-uPAR complex and inhibiting its signaling and internalization.Am J Physiol Cell Physiol. 2008 Jul;295(1):C257-67. doi: 10.1152/ajpcell.00569.2007. Epub 2008 May 21. Am J Physiol Cell Physiol. 2008. PMID: 18495808 Free PMC article.
-
Endothelial cell dysfunction and cytoskeletal changes associated with repression of p16(INK4a) during immortalization.Oncogene. 2012 Nov 15;31(46):4815-27. doi: 10.1038/onc.2011.645. Epub 2012 Feb 6. Oncogene. 2012. PMID: 22310292 Free PMC article.
-
Kininogen enhances seizure susceptibility in mice possibly through bradykinin-induced modulation of calcium transients in glutamatergic and GABAergic neurons.Front Pharmacol. 2025 Jun 10;16:1509837. doi: 10.3389/fphar.2025.1509837. eCollection 2025. Front Pharmacol. 2025. PMID: 40556759 Free PMC article.
References
-
- Folkman J. (1995) Nat. Med. 1, 27-31. - PubMed
-
- Folkman J. (1985) Adv. Cancer Res. 43, 175-203. - PubMed
-
- Shweiki D., Itin, A., Soffer, D. & Keshet, E. (1992) Nature (London) 359, 843-845. - PubMed
-
- Thomas K. A. (1996) J. Biol. Chem. 271, 603-606. - PubMed
-
- Wang Y. & Becker, D. (1997) Nat. Med. 3, 887-893. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
