

Interaction between domains of a plant NBS-LRR protein in disease resistance-related cell death
- PMID: 12198153
- PMCID: PMC126192
- DOI: 10.1093/emboj/cdf453
Interaction between domains of a plant NBS-LRR protein in disease resistance-related cell death
Abstract
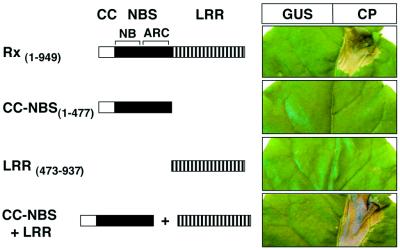
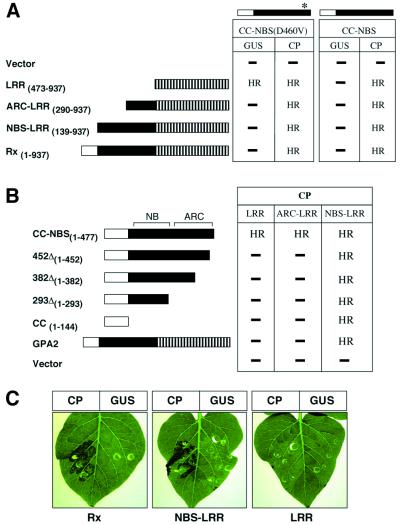
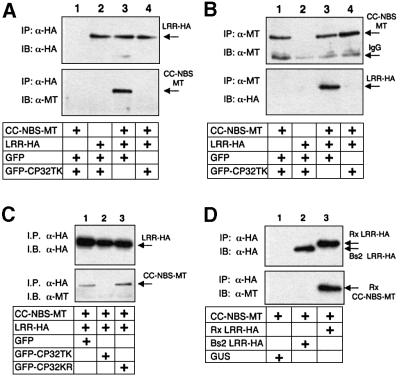
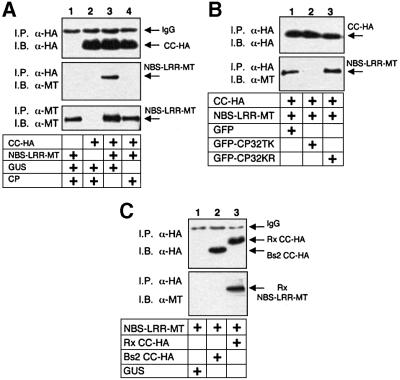
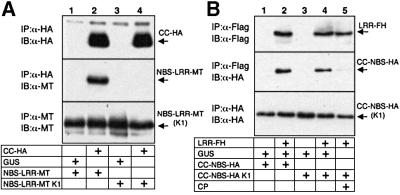
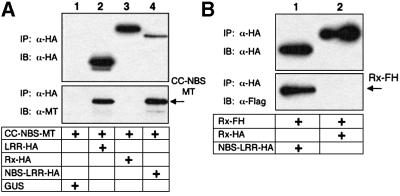
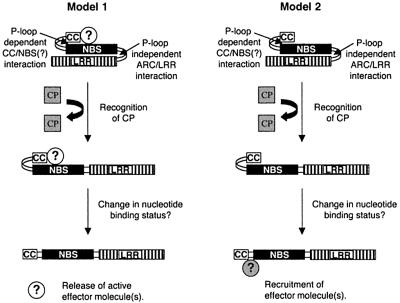
Many plant disease resistance (R) genes encode proteins predicted to have an N-terminal coiled-coil (CC) domain, a central nucleotide-binding site (NBS) domain and a C-terminal leucine-rich repeat (LRR) domain. These CC-NBS-LRR proteins recognize specific pathogen-derived products and initiate a resistance response that often includes a type of cell death known as the hypersensitive response (HR). Co-expression of the potato CC-NBS-LRR protein Rx and its elicitor, the PVX coat protein (CP), results in a rapid HR. Surprisingly, co-expression of the LRR and CC-NBS as separate domains also resulted in a CP-dependent HR. Likewise, the CC domain complemented a version of Rx lacking this domain (NBS- LRR). Correspondingly, the LRR domain interacted physically in planta with the CC-NBS, as did CC with NBS-LRR. Both interactions were disrupted in the presence of CP. However, the interaction between CC and NBS-LRR was dependent on a wild-type P-loop motif, whereas the interaction between CC-NBS and LRR was not. We propose that activation of Rx entails sequential disruption of at least two intramolecular interactions.
Figures
References
-
- Aravind L., Dixit,V.M. and Koonin,E.V. (1999) The domains of death: evolution of the apoptosis machinery. Trends Biochem. Sci., 24, 47–53. - PubMed
-
- Austin M.J., Muskett,P., Kahn,K., Feys,B.J., Jones,J.D.G. and Parker,J.E. (2002) Regulatory role of SGT1 in early R gene-mediated plant defenses. Science, 295, 2077–2080. - PubMed
-
- Ayliffe M.A., Frost,D.V., Finnegan,E.J., Lawrence,G.J., Anderson,P.A. and Ellis,J.G. (1999) Analysis of alternative transcripts of the flax L6 rust resistance gene. Plant J., 17, 287–292. - PubMed
-
- Azevedo C., Sadanandom,A., Kitagawa,K., Freialdenhoven,A., Shirasu,K. and Schulze-Lefert,P. (2002) The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science, 295, 2073–2076. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
