

The chromatin remodeling complex NoRC targets HDAC1 to the ribosomal gene promoter and represses RNA polymerase I transcription
- PMID: 12198165
- PMCID: PMC126197
- DOI: 10.1093/emboj/cdf460
The chromatin remodeling complex NoRC targets HDAC1 to the ribosomal gene promoter and represses RNA polymerase I transcription
Abstract
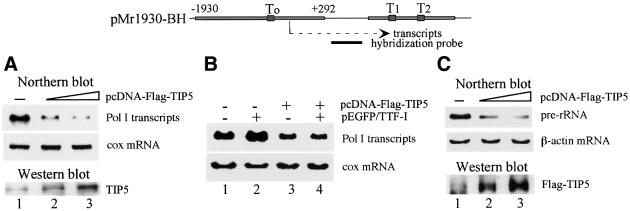
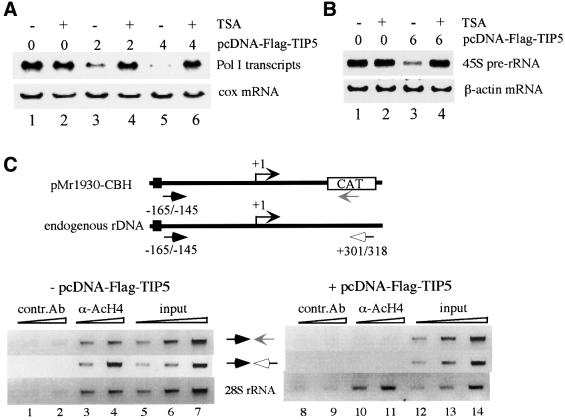
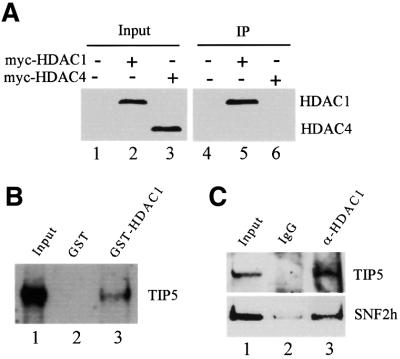
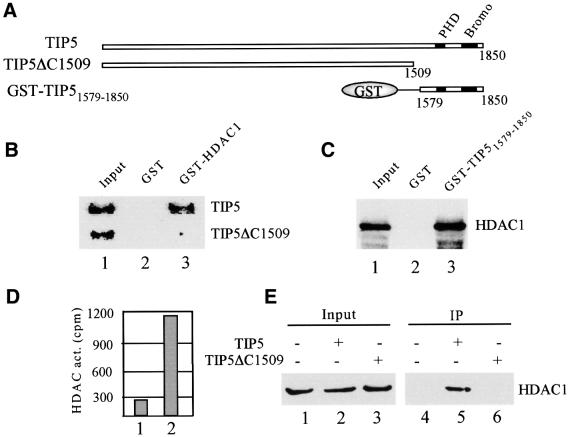
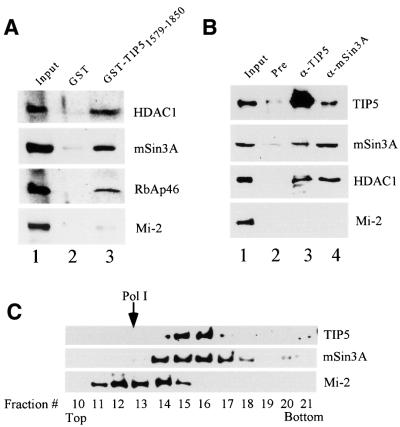
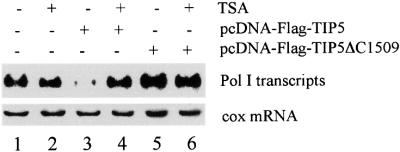
Mammalian chromatin remodeling complexes are involved in both activation and repression of transcription. Here, we show that NoRC, a SNF2h- containing nucleolar chromatin remodeling complex, represses ribosomal gene transcription. NoRC-mediated rDNA silencing was alleviated by trichostatin A, indicating that histone deacetylation is causally involved in silencing. Chromatin immunoprecipitation experiments demonstrate that overexpression of TIP5, the large subunit of NoRC, mediates deacetylation of nucleosomes in the vicinity of the rDNA promoter. Protein-protein interaction assays reveal association of TIP5 with the histone deacetylase HDAC1 in vivo and in vitro. Deletion of the C-terminal PHD finger and bromodomain abolishes the interaction of TIP5 and HDAC1, and abrogates transcriptional repression. The results suggest that NoRC silences the rDNA locus by targeting the SIN3 corepressor complex to the rDNA promoter, thereby establishing a repressed chromatin structure.
Figures
References
-
- Ahringer J. (2000) NuRD and SIN3 histone deacetylase complexes in development. Trends Genet., 16, 351–356. - PubMed
-
- Ayer D. (1999) Histone deacetylases: transcriptional repression with SINers and NuRDs. Trends Cell Biol., 9, 193–198. - PubMed
-
- Brown C.J. and Robinson,W.P. (2000) The causes and consequences of random and non-random X chromosome inactivation in humans. Clin. Genet., 58, 353–363. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
