

Telomere formation on macronuclear chromosomes of Oxytricha trifallax and O. fallax: alternatively processed regions have multiple telomere addition sites
- PMID: 12199911
- PMCID: PMC128808
- DOI: 10.1186/1471-2156-3-16
Telomere formation on macronuclear chromosomes of Oxytricha trifallax and O. fallax: alternatively processed regions have multiple telomere addition sites
Abstract
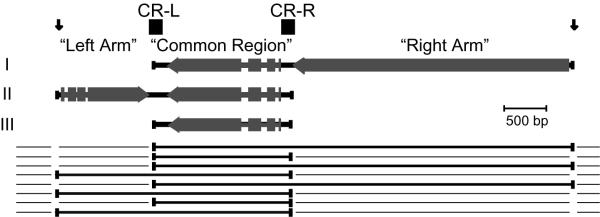
Background: Ciliates employ massive chromatid breakage and de novo telomere formation during generation of the somatic macronucleus. Positions flanking the 81-MAC locus are reproducibly cut. But those flanking the Common Region are proposed to often escape cutting, generating three nested macronuclear chromosomes, two retaining "arms" still appended to the Common Region. Arm-distal positions must differ (in cis) from the Common Region flanks.
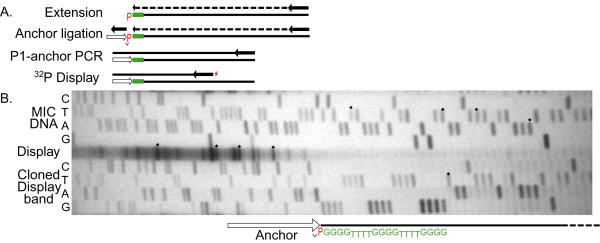
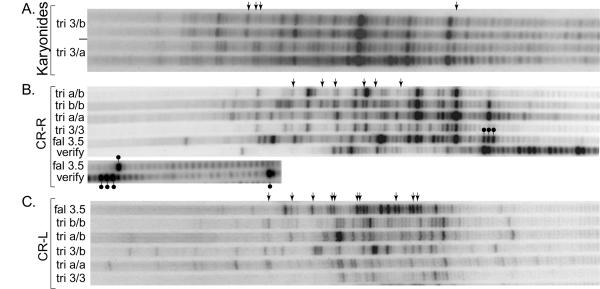
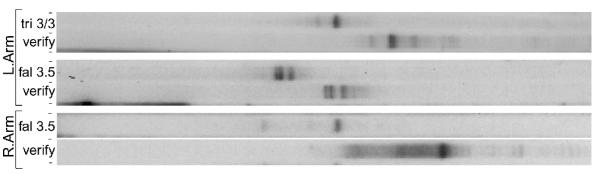
Results: The Common-Region-flanking positions also differ from the arm-distal positions in that they are "multi-TAS" regions: anchored PCR shows heterogeneous patterns of telomere addition sites, but arm-distal sites do not. The multi-TAS patterns are reproducible, but are sensitive to the sequence of the allele being processed. Thus, random degradation following chromatid cutting does not create this heterogeneity; these telomere addition sites also must be dictated by cis-acting sequences.
Conclusions: Most ciliates show such micro-heterogeneity in the precise positions of telomere addition sites. Telomerase is believed to be tightly associated with, and act in concert with, the chromatid-cutting nuclease: heterogeneity must be the result of intervening erosion activity. Our "weak-sites" hypothesis explains the correlation between alternative chromatid cutting at the Common Region boundaries and their multi-TAS character: when the chromatid-breakage machine encounters either a weak binding site or a weak cut site at these regions, then telomerase dissociates prematurely, leaving the new end subject to erosion by an exonuclease, which pauses at cis-acting sequences; telomerase eventually heals these resected termini. Finally, we observe TAS positioning influenced by trans-allelic interactions, reminiscent of transvection.
Figures

Similar articles
-
Two two-gene macronuclear chromosomes of the hypotrichous ciliates Oxytricha fallax and O. trifallax generated by alternative processing of the 81 locus.Dev Genet. 1997;20(4):348-57. doi: 10.1002/(SICI)1520-6408(1997)20:4<348::AID-DVG6>3.0.CO;2-7. Dev Genet. 1997. PMID: 9254909
-
Developmental precise excision of Oxytricha trifallax telomere-bearing elements and formation of circles closed by a copy of the flanking target duplication.EMBO J. 1993 Dec;12(12):4593-601. doi: 10.1002/j.1460-2075.1993.tb06148.x. EMBO J. 1993. PMID: 8223469 Free PMC article.
-
The Oxytricha trifallax macronuclear genome: a complex eukaryotic genome with 16,000 tiny chromosomes.PLoS Biol. 2013;11(1):e1001473. doi: 10.1371/journal.pbio.1001473. Epub 2013 Jan 29. PLoS Biol. 2013. PMID: 23382650 Free PMC article.
-
Multiple sequence versions of the Oxytricha fallax 81-MAC alternate processing family.J Protozool. 1987 Nov;34(4):429-34. doi: 10.1111/j.1550-7408.1987.tb03207.x. J Protozool. 1987. PMID: 3123649 Review.
-
Chromosome healing by de novo telomere addition in Saccharomyces cerevisiae.Mol Microbiol. 2006 Mar;59(5):1357-68. doi: 10.1111/j.1365-2958.2006.05026.x. Mol Microbiol. 2006. PMID: 16468981 Review.
Cited by
-
Programmed chromosome fragmentation in ciliated protozoa: multiple means to chromosome ends.Microbiol Mol Biol Rev. 2023 Dec 20;87(4):e0018422. doi: 10.1128/mmbr.00184-22. Epub 2023 Nov 27. Microbiol Mol Biol Rev. 2023. PMID: 38009915 Free PMC article. Review.
-
Chromosome fusions triggered by noncoding RNA.RNA Biol. 2017 May 4;14(5):620-631. doi: 10.1080/15476286.2016.1195940. Epub 2016 Jun 7. RNA Biol. 2017. PMID: 27267579 Free PMC article.
-
Programmed Genome Rearrangements in the Ciliate Oxytricha.Microbiol Spectr. 2014 Dec;2(6):10.1128/microbiolspec.MDNA3-0025-2014. doi: 10.1128/microbiolspec.MDNA3-0025-2014. Microbiol Spectr. 2014. PMID: 26104449 Free PMC article. Review.
-
Programmed genome rearrangements in ciliates.Cell Mol Life Sci. 2020 Nov;77(22):4615-4629. doi: 10.1007/s00018-020-03555-2. Epub 2020 May 27. Cell Mol Life Sci. 2020. PMID: 32462406 Free PMC article. Review.
-
A chimeric chromosome in the ciliate oxytricha resulting from duplication.J Mol Evol. 2011 Oct;73(3-4):70-3. doi: 10.1007/s00239-011-9464-1. Epub 2011 Oct 22. J Mol Evol. 2011. PMID: 22037698 Free PMC article.
References
-
- B McClintock. The fusion of broken ends of sister half-chromatids following chromatid breakage at meiotic anaphases. Missouri Agricultural Experiment Station Research Bulletin. 1938;290:1–48.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
