

A gene mutated in nephronophthisis and retinitis pigmentosa encodes a novel protein, nephroretinin, conserved in evolution
- PMID: 12205563
- PMCID: PMC385091
- DOI: 10.1086/344395
A gene mutated in nephronophthisis and retinitis pigmentosa encodes a novel protein, nephroretinin, conserved in evolution
Abstract
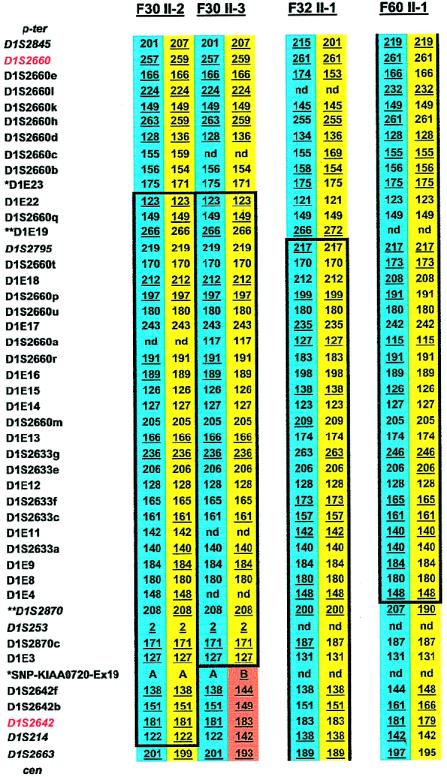
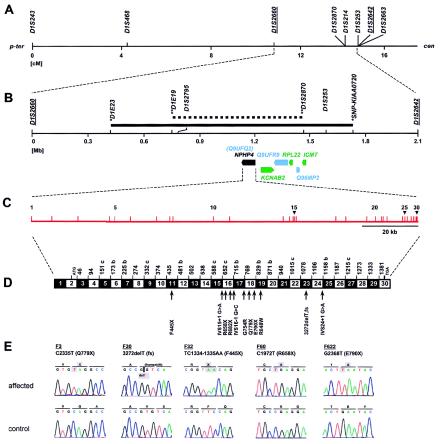
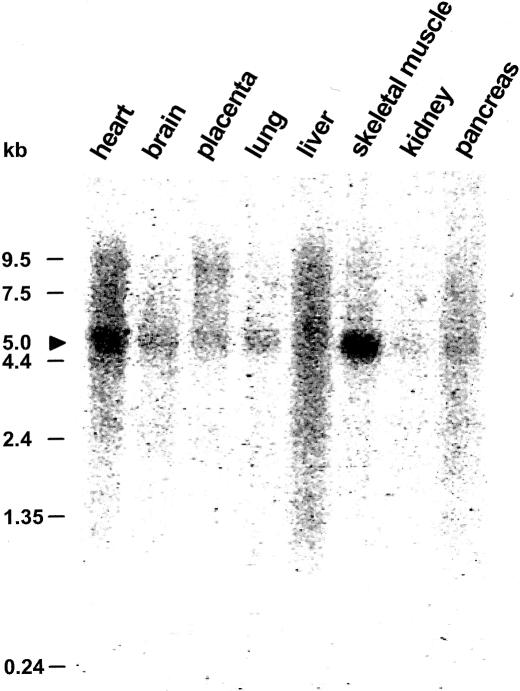
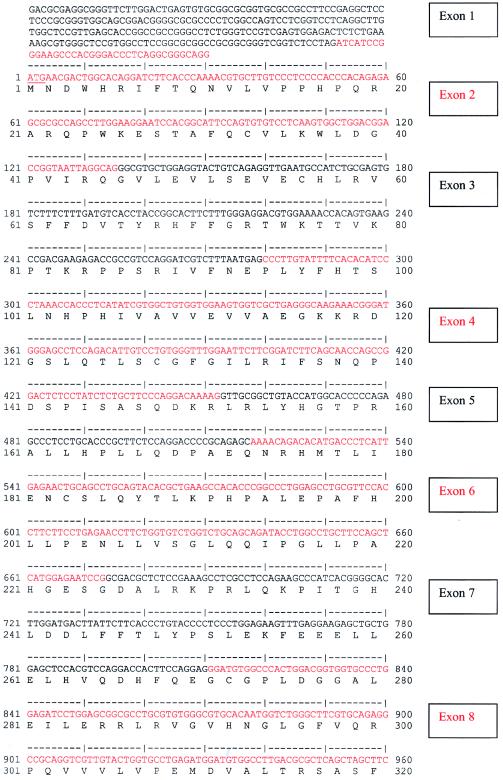
Nephronophthisis (NPHP) comprises a group of autosomal recessive cystic kidney diseases, which constitute the most frequent genetic cause for end-stage renal failure in children and young adults. The most prominent histologic feature of NPHP consists of development of renal fibrosis, which, in chronic renal failure of any origin, represents the pathogenic event correlated most strongly to loss of renal function. Four gene loci for NPHP have been mapped to chromosomes 2q13 (NPHP1), 9q22 (NPHP2), 3q22 (NPHP3), and 1p36 (NPHP4). At all four loci, linkage has also been demonstrated in families with the association of NPHP and retinitis pigmentosa, known as "Senior-Løken syndrome" (SLS). Identification of the gene for NPHP type 1 had revealed nephrocystin as a novel docking protein, providing new insights into mechanisms of cell-cell and cell-matrix signaling. We here report identification of the gene (NPHP4) causing NPHP type 4, by use of high-resolution haplotype analysis and by demonstration of nine likely loss-of-function mutations in six affected families. NPHP4 encodes a novel protein, nephroretinin, that is conserved in evolution--for example, in the nematode Caenorhabditis elegans. In addition, we demonstrate two loss-of-function mutations of NPHP4 in patients from two families with SLS. Thus, we have identified a novel gene with critical roles in renal tissue architecture and ophthalmic function.
Figures
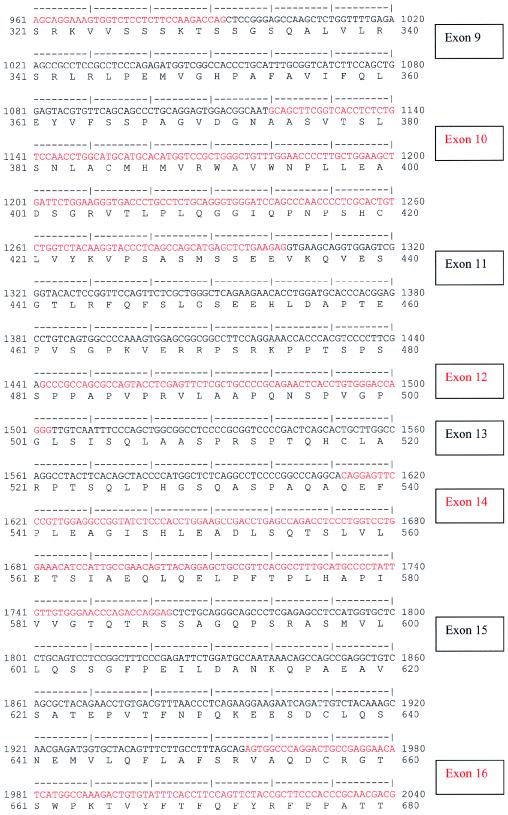
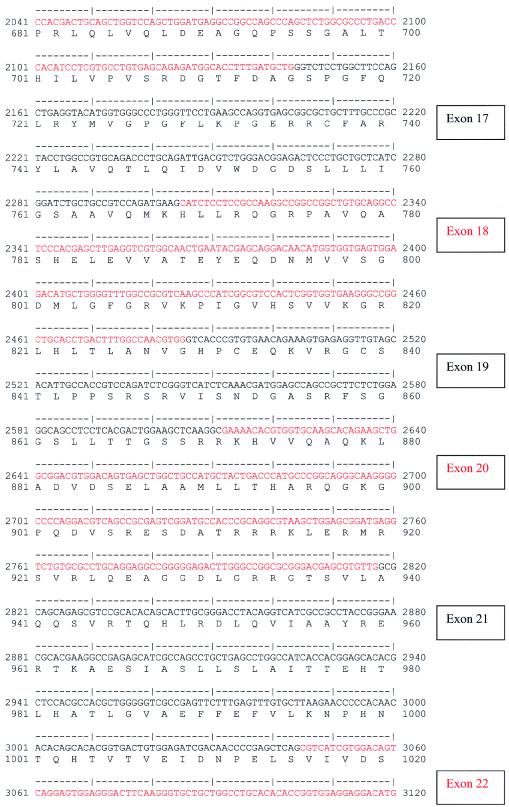
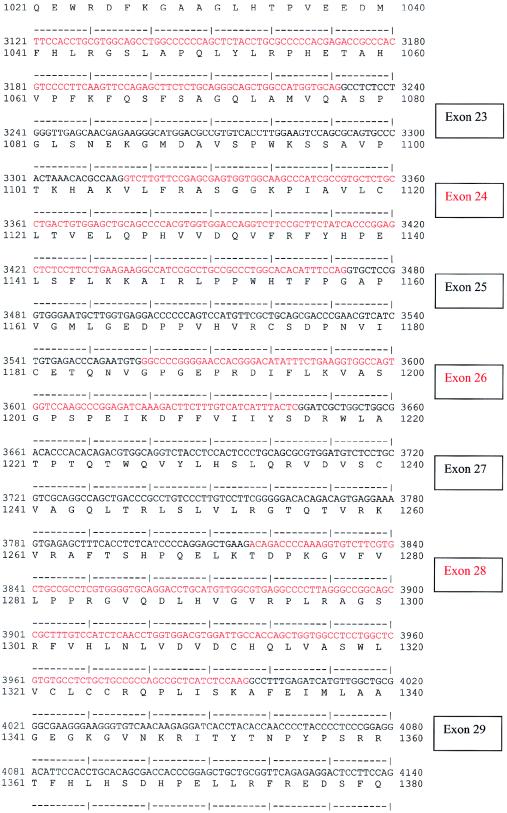
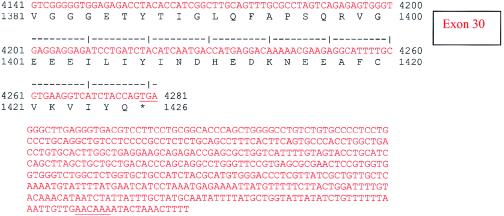


References
Electronic-Database Information
-
- AceView, http://www.ncbi.nlm.nih.gov/IEB/Research/Acembly/ (for genomic structure)
-
- Ensembl Genome Browser, http://www.ensembl.org/ (for genomic structure of NPHP4)
-
- ExPASy Molecular Biology Server, http://www.expasy.ch/ (for Tmpred)
-
- Hits Home, http://hits.isb-sib.ch/ (for MOTIF SCAN program)
-
- Marshfield Medical Research Foundation Home Page, http://research.marshfieldclinic.org/ (for map of microsatellite markers)
References
-
- Antignac C, Arduy C, Beckmann JS, Benessy F, Gros F, Medhioub M, Hildebrandt F, Dufier JL, Kleinknecht C, Broyer M (1993) A gene for familial juvenile nephronophthisis (recessive medullary cystic disease) maps to chromosome 2p. Nat Genet 3:342–345 - PubMed
-
- Caridi G, Murer L, Bellantuono R, Sorino P, Caringella DA, Gusmano R, Ghiggeri GM (1998) Renal-retinal syndromes: association of retinal anomalies and recessive nephronophthisis in patients with homozygous deletion of the NPH1 locus. Am J Kidney Dis 32:1059–1062 - PubMed
-
- Dib C, Faure S, Fizames C, Samson D, Drouot N, Vignal A, Millasseau P, Hazan J, Seboun E, Lathrop M, Gyapay G, Morissette J, Weissenbach, J (1996) A comprehensive genetic map of the human genome based on 5,264 microsatellites. Nature 380:152–154 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
