

doi: 10.1101/gad.230702.
Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells
Affiliations
- PMID: 12208843
- PMCID: PMC186665
- DOI: 10.1101/gad.230702
Item in Clipboard
Shoot meristem maintenance is controlled by a GRAS-gene mediated signal from differentiating cells
Genes Dev.
.
Abstract
Plant shoot development depends on the perpetuation of a group of undifferentiated cells in the shoot apical meristem (SAM). In the Petunia mutant hairy meristem (ham), shoot meristems differentiate postembryonically as continuations of the subtending stem. HAM encodes a putative transcription factor of the GRAS family, which acts non-cell-autonomously from L3-derived tissue of lateral organ primordia and stem provasculature. HAM acts in parallel with TERMINATOR (PhWUSCHEL) and is required for continued cellular response to TERMINATOR and SHOOTMERISTEMLESS (PhSTM). This reveals a novel mechanism by which signals from differentiating tissues extrinsically control stem cell fate in the shoot apex.
Figures

Phenotype of ham mutants. (A)
Wild-type W138 Petunia. (B) Wild-type W138 flower
with the internal whorls of organs. Five stamens (yellow arrow)
surround two fused carpels (green arrow). (C)
ham-B4281 plant, terminated during vegetative rosette growth.
(D) ham mutant flower lacking two stamens and both
carpels. (E) Wild-type vegetative apex. p3, p2, and
p1 = leaf primordia in order of decreasing age; m = meristem.
(F) Vegetative ham apex shortly after termination.
Primordium initiation has ceased. The meristem displays ectopic
trichomes (arrows). (G) As in F, 2 wk after
termination, the central dome has increased in size and is covered with
trichomes. (H) Wild-type inflorescence apex. fm = floral
meristem, im = inflorescence meristem, br = bract, se = sepal.
(I) ham inflorescence apex with ectopic trichomes.
br = last initiated bracts. (J) ham floral
meristem, showing termination after initiation of three stamens (st).
In place of carpels, a flat apex is visible with a small outgrowth
(arrow). Bars, 100 μm.
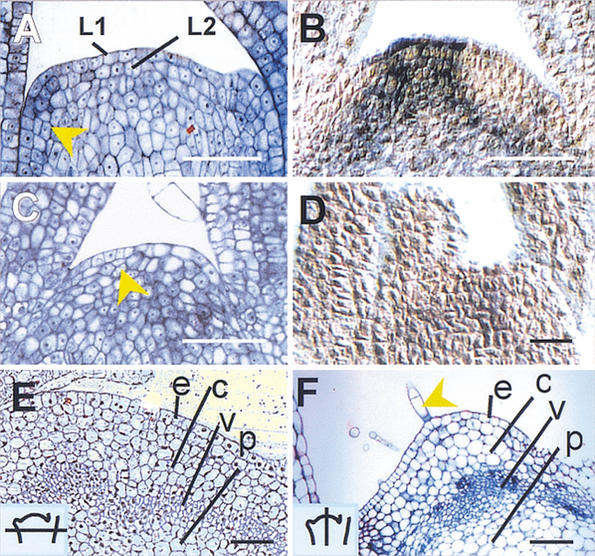
Histology of ham apices. (A)
Wild-type vegetative meristem. The arrow indicates a periclinal
division in the L2 layer of an initiating leaf primordium. (B)
In situ localization of PhSTM transcript in a wild-type
vegetative apex. The signal (blue) is excluded from the leaf primordia.
(C) ham vegetative apex showing a cessation of organ
initiation and periclinal division in the central zone (arrow).
(D) In situ localization of PhSTM transcript in a
ham apex shortly after termination. (E) Transverse
section of developing stem, just below a wild-type meristem (section
schematized, inset). e = epidermis, c = cortex,
v = vasculature, p = pith. (F) Older ham apex in
longitudinal section showing a layered structure of differentiated
tissue. e, c, v, p as in E. Arrow = trichome. Bars, 50
μm.
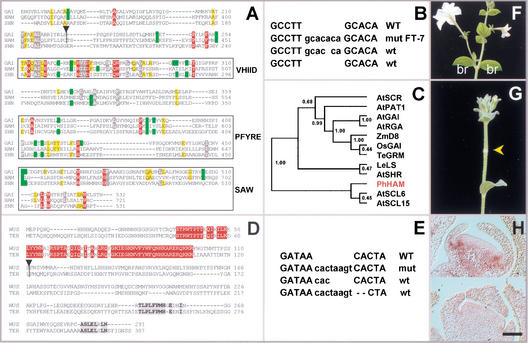
Cloning and structure of HAM and
TER. (A) Protein sequence alignment of the C-terminal
portion of HAM with SHR and GAI. Absolutely
conserved positions are red, and conserved residues are grey (>90%),
yellow (>80%), or green (>70%), based on alignments of 12 GRAS
sequences as in the cladogram of C. Atypical residues at
conserved positions are not colored. VHIID, PFYRE, and SAW are domains
as defined in Pysh et al. (1999). Triangle = dTph1 insertion
in ham-B4281. (B) ham excision alleles.
WT = wild-type sequence flanking the insertion in ham-B4281,
wt = footprints restoring HAM function, mut = mutant
footprint allele hamFT-7. (C) Tree produced by
neighbor joining (ClustalG software) showing the similarities of 12
GRAS sequences. Numbers indicate bootstrap frequencies of each
branchpoint in the cladogram. GenBank accession nos. AtGAI (CAA75492),
AtSHR (NP195480), AtPAT1 (AAF73237), AtRGA (CAA75493), LeLS (AAD05242),
AtSCL6 (NP191926), AtSCR (AAB06318), AtSCL15 (NP191622), ZmD8
(AAL10319), TeGRM (CAB51555), OsGAI (BAA90749), and PhHAM (AF481952).
(D) Full protein alignment of TER (GenBank accession no.
AF481951) and WUS (GenBank accession no. CAA09986). Conservation is
given on the basis of these two orthologs only. Red residues indicate
positions in the homeobox, and grey residues denote blocks of
conspicuous colinearity. (E) As in B but for
ter-B1382. (F) Wild-type Petunia
inflorescence producing two bracts (br) and a flower per node.
(G) HAM cosuppressor showing a node without bracts
and flower (arrow). (H) In situ hybridization of HAM
RNA on wild-type (upper) and cosuppressed (lower)
floral meristems. Wild type shows a signal in the initiating petal
primordia. The cosuppressor lacks this signal. Bar, 50 μm.
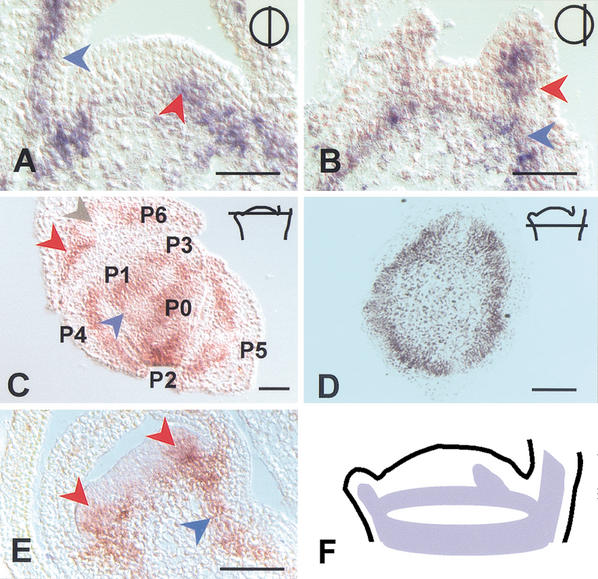
Expression pattern of HAM. (A) In
situ localization of HAM transcript in a near median (top
right inset) longitudinal section through a vegetative apex. Signal
is in the developing primordia (blue arrow) and at the presumptive
position of a newly initiating primordium (red arrow). (B) As
in A, with a section located more peripherally (top
right inset). The signal is seen in a developing primordium (red
arrow), as well as in a ring-shaped pattern that corresponds to the
developing stem vasculature (blue arrow). (C) As in
A, but in a transverse section. The position of the section is
indicated in the top right inset. The signal is observed in
the inner ground tissues of the primordia (red arrow) and is weaker in
the main vascular bundle of older primordia (grey arrow). The blue
arrow indicates HAM expression in a ring-shaped pattern that
merges with primordia P1 and P0 and corresponds to provascular tissue
of the differentiating stem. P6, P5, and so forth indicate the
consecutive order of primordium initiation with decreasing age.
(D) As in C, but at a position just below the
meristem (indicated in top right inset). HAM
expression is seen as a ring that corresponds to the provasculature of
the stem. (E) HAM localization during development of
the floral meristem, as exemplified for initiating petal primordia.
Expression is observed in inner cell layers at the site of petal
initiation (red arrows) and in subtending provascular tissue of the
developing pedicel (blue arrow). (F) Schematic representation
of the HAM expression pattern as exemplified for a wild-type
vegetative meristem. Bar, 50 μm.
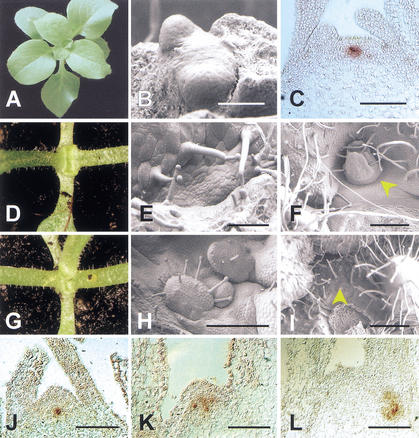
Relations between ham and ter.
(A) Wild-type Petunia during vegetative rosette
growth. (B) Wild-type shoot apical meristem (SAM) with the
first two true leaf primordia. Cotyledons have been removed.
(C) In situ localization of TER (PhWUS)
transcripts in a wild-type vegetative apex. (D)
ter-B1382 seedling. Growth has ceased after production of the
first two true leaves. (E) ter seedling apex after
initiation of the two first leaves. A flat, disorganized structure
replaces the SAM. (F) ter apex with an ectopic
meristem (arrow, stop-and-go growth). (G) ter ham
double mutant seedling. An additional leaf, compared with D,
occurred with a low frequency in ter single mutants as well.
(H) ter ham double mutant seedling apex. Initiation
of ectopic leaves is observed (stop-and-go). (I) ter
ham double mutant apex on an older plant. The SAM displays a
trichome covered surface characteristic for ham single
mutants. (J) In situ localization of PhWUS
transcripts in a ham mutant apex shortly after termination.
The signal is essentially normal. (K) In situ localization of
PhWUS transcripts in a later ham mutant apex.
Expression occurs in a disorganized pattern. (L) As in
K. PhWUS expression in the main apex has disappeared.
In the axillary position, expression is disorganized and deeply
internal. Bars: B, 25 μm; C,E,J, 50 μm;
F,I, 200 μm; H,K,L, 100 μm.
References
-
- Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R. Dependence of stem cell fate in Arabidopsison a feedback loop regulated by CLV3 activity. Science. 2000;289:617–619. - PubMed
-
- Byrne M, Barley R, Curtis M, Arroyo JM, Dunham M, Hudson A, Martienssen RA. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature. 2000;408:967–971. - PubMed
-
- Janssen BJ, Williams A, Chen JJ, Mathern J, Hake S, Sinha N. Isolation and characterisation of two knotted-like homeobox genes from tomato. Plant Mol Biol. 1998;36:417–425. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous