

Cancer predisposition and hematopoietic failure in Rad50(S/S) mice
- PMID: 12208847
- PMCID: PMC186667
- DOI: 10.1101/gad.1007902
Cancer predisposition and hematopoietic failure in Rad50(S/S) mice
Abstract
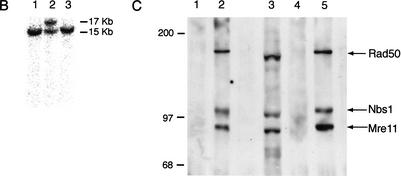
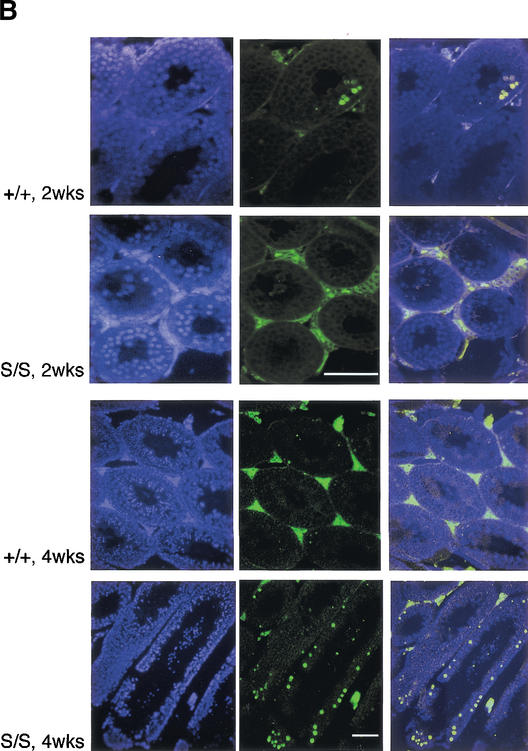
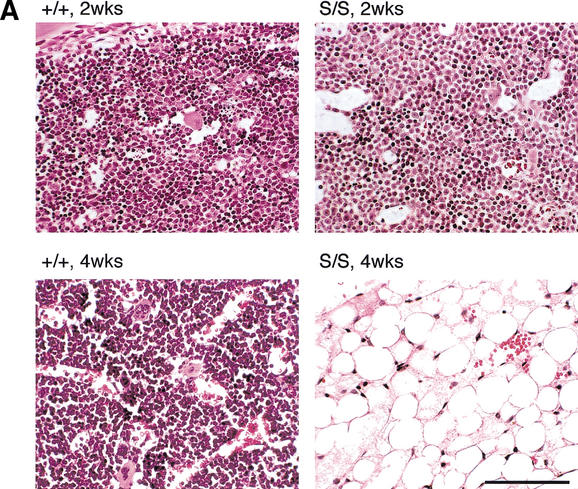
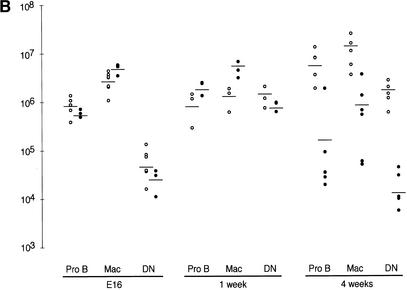
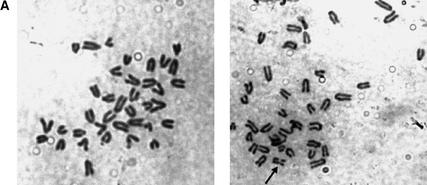
Mre11, Rad50, and Nbs1 function in a protein complex that is central to the metabolism of chromosome breaks. Null mutants of each are inviable. We demonstrate here that hypomorphic Rad50 mutant mice (Rad50(S/S) mice) exhibited growth defects and cancer predisposition. Rad50(S/S) mice died with complete bone marrow depletion as a result of progressive hematopoietic stem cell failure. Similar attrition occurred in spermatogenic cells. In both contexts, attrition was substantially mitigated by p53 deficiency, whereas the tumor latency of p53(-/-) and p53(+/-) animals was reduced by Rad50(S/S). Indices of genotoxic stress and chromosomal rearrangements were evident in Rad50(S/S) cultured cells, as well as in Rad50(S/S) and p53(-/-) Rad50(S/S) lymphomas, suggesting that the Rad50(S/S) phenotype was attributable to chromosomal instability. These outcomes were not associated with overt defects in the Mre11 complex's previously established double strand break repair and cell cycle checkpoint regulation functions. The data indicate that even subtle perturbation of Mre11 complex functions results in severe genotoxic stress, and that the complex is critically important for homeostasis of proliferative tissues.
Figures










References
-
- Alani E, Padmore R, Kleckner N. Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell. 1990;61:419–436. - PubMed
-
- Baudat F, Manova K, Yuen JP, Jasin M, Keeney S. Chromosome synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol Cell. 2000;6:989–998. - PubMed
-
- Carney JP, Maser RS, Olivares H, Davis EM, Le Beau M, Yates JR, 3rd, Hays L, Morgan WF, Petrini JHJ. The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: Linkage of double-strand break repair to the cellular DNA damage response. Cell. 1998;93:477–486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous