

An ATF2-derived peptide sensitizes melanomas to apoptosis and inhibits their growth and metastasis
- PMID: 12208865
- PMCID: PMC151112
- DOI: 10.1172/JCI16081
An ATF2-derived peptide sensitizes melanomas to apoptosis and inhibits their growth and metastasis
Abstract
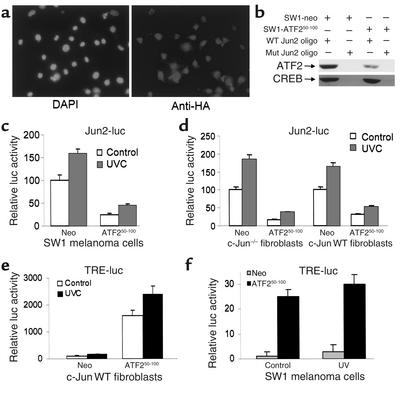
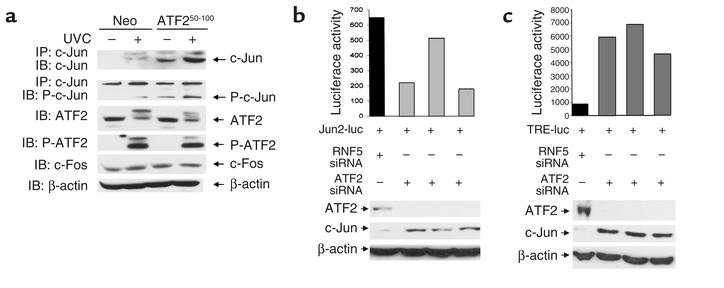
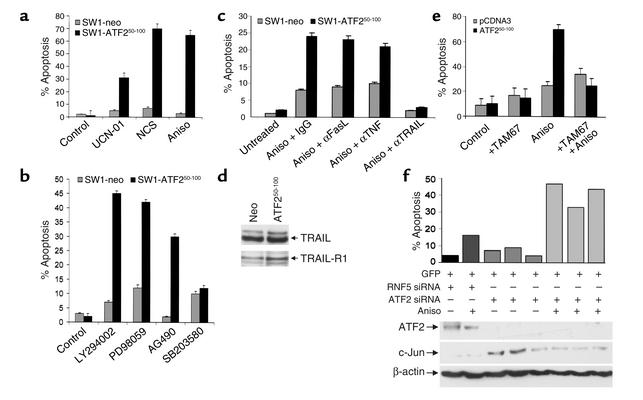
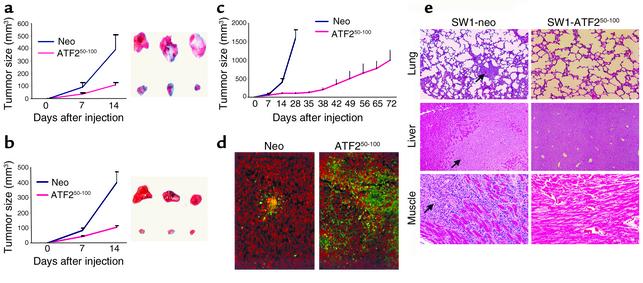
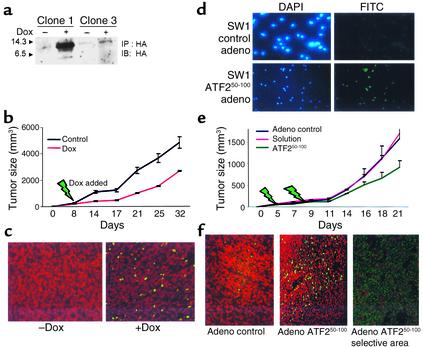
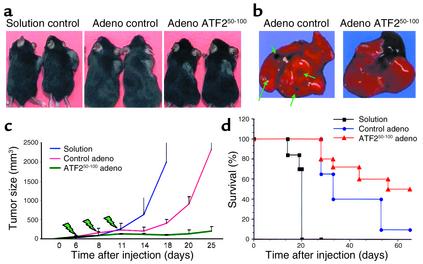
Melanomas are among the aggressive tumor types because of their notorious resistance to treatment and their high capacity to metastasize. ATF2 is among transcription factors implicated in the progression of melanoma and its resistance to treatment. Here we demonstrate that the expression of a peptide spanning amino acids 50-100 of ATF2 (ATF2(50-100)) reduces ATF2 transcriptional activities while increasing the expression and activity of c-Jun. Altering the balance of Jun/ATF2 transcriptional activities sensitized melanoma cells to apoptosis, an effect that could be attenuated by inhibiting c-Jun. Inhibition of ATF2 via RNA interference likewise increased c-Jun expression and primed melanoma cells to undergo apoptosis. Growth and metastasis of SW1 and B16F10 mouse melanomas were inhibited by ATF2(50-100) to varying degrees up to a complete regression, depending on the mode (inducible, constitutive, or adenoviral delivery) of its expression.
Figures
References
-
- Yang YM, Dolan LR, Ronai Z. Expression of dominant negative CREB reduces resistance to radiation of human melanoma cells. Oncogene. 1996;12:2223–2233. - PubMed
-
- Jean D, Harbison M, McConkey DJ, Ronai Z, Bar-Eli M. CREB and its associated proteins act as survival factors for human melanoma cells. J. Biol Chem. 1998;273:24884–24890. - PubMed
-
- Yang J, Richmond A. Constitutive IkappaB kinase activity correlates with nuclear factor-kappaB activation in human melanoma cells. Cancer Res. 2001;61:4901–4909. - PubMed
-
- Recio JA, Merlino G. Hepatocyte growth factor/scatter factor activates proliferation in melanoma cells through p38 MAPK, ATF-2 and cyclin D1. Oncogene. 2002;21:1000–1008. - PubMed
-
- Ronai Z, et al. ATF2 confers radiation resistance to human melanoma cells. Oncogene. 1998;16:523–531. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
