

The mengovirus leader protein suppresses alpha/beta interferon production by inhibition of the iron/ferritin-mediated activation of NF-kappa B
- PMID: 12208945
- PMCID: PMC136484
- DOI: 10.1128/jvi.76.19.9664-9672.2002
The mengovirus leader protein suppresses alpha/beta interferon production by inhibition of the iron/ferritin-mediated activation of NF-kappa B
Abstract
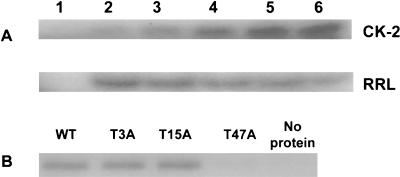
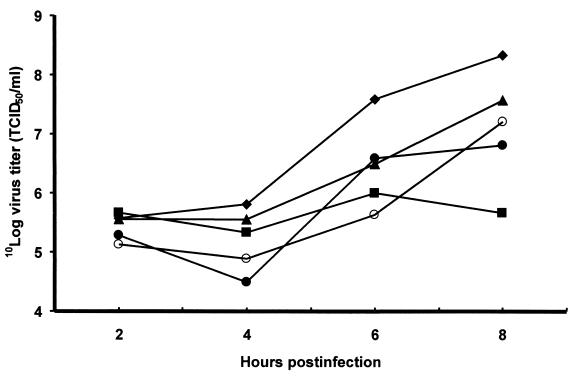
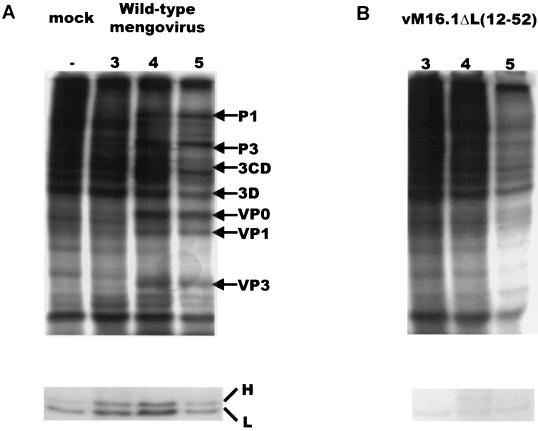
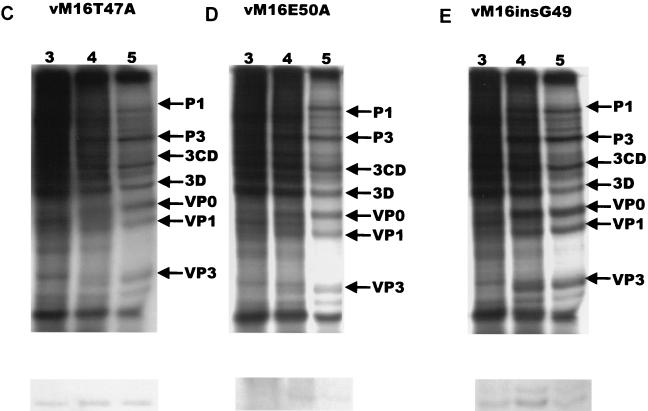
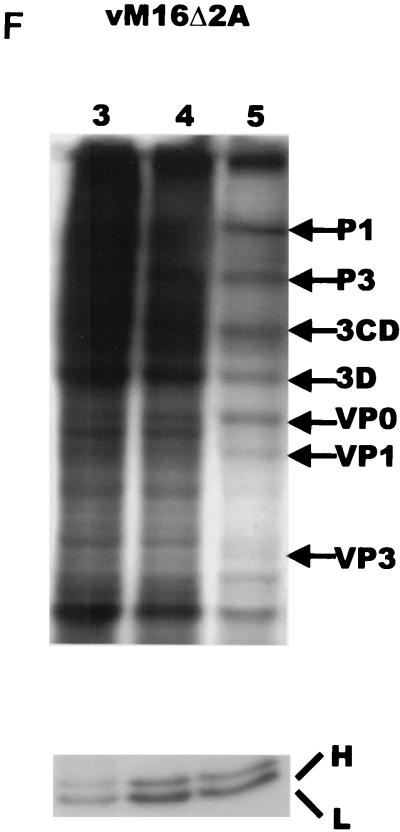
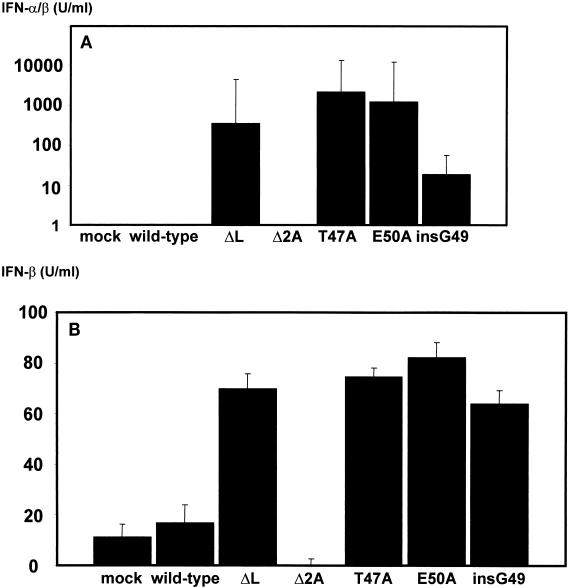
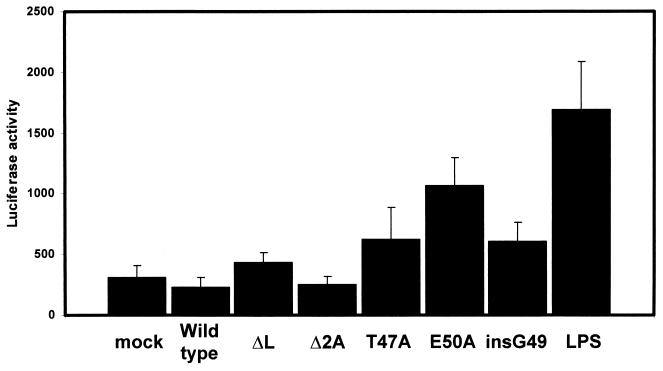
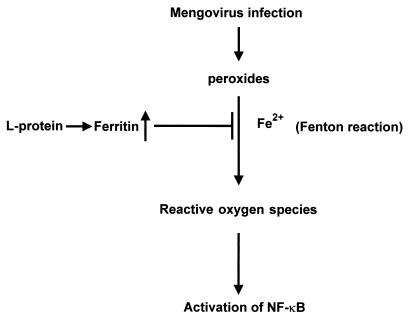
In our studies on the biological function of the mengovirus leader protein, we identified a casein kinase II (CK-2) phosphorylation site in the protein. Here we report that the mengovirus leader protein can be phosphorylated by CK-2 in vitro. Expression of a recombinant leader protein in which the consensus CK-2 sequence around threonine 47 was disturbed resulted in a mutant protein that could no longer be phosphorylated. The CK-2 consensus sequence was modified by site-directed mutagenesis and subsequently introduced into a mengovirus cDNA clone to investigate the effect of the phosphorylation of the leader protein on virus replication and on the host cell response. Modifications by which the CK-2 consensus sequence was disturbed resulted in mutant viruses with reduced growth kinetics. We demonstrated that the integrity of the CK-2 phosphorylation site of the mengovirus leader protein was specifically related to the suppression of NF-kappa B activation and subsequent suppression of alpha/beta interferon production in infected cells. We also found that the integrity of the CK-2 phosphorylation site of the leader protein coincided with an increase of ferritin expression in the infected cell. These data indicate that the leader protein suppresses the iron-mediated activation of NF-kappa B and thereby inhibits alpha/beta interferon expression in the infected cell.
Figures
References
-
- Aisen, P., C. Enns, and M. Wessling-Resnick. 2001. Chemistry and biology of eukaryotic iron metabolism. Int. J. Biochem. Cell. Biol. 33:940-959. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. A. Smith, J. G. Seidman, and K. Struhl. 1995. Current protocols in molecular biology. John Wiley & Sons, New York, N.Y.
-
- Boege, U., R. Hancharyk, and D. G. Scraba. 1987. The synthesis of a particle-forming cellular protein is enhanced by Mengo virus infection. Virology 159:358-367. - PubMed
-
- Cairo, G., L. Tacchini, G. Pogliaghi, E. Anzon, A. Tomasi, and A. Bernelli-Zazzera. 1995. Induction of ferritin synthesis by oxidative stress. Transcriptional and post-transcriptional regulation by expansion of the “free” iron pool. J. Biol. Chem. 270:700-703. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
