

Strategy for nonenveloped virus entry: a hydrophobic conformer of the reovirus membrane penetration protein micro 1 mediates membrane disruption
- PMID: 12208969
- PMCID: PMC136509
- DOI: 10.1128/jvi.76.19.9920-9933.2002
Strategy for nonenveloped virus entry: a hydrophobic conformer of the reovirus membrane penetration protein micro 1 mediates membrane disruption
Abstract
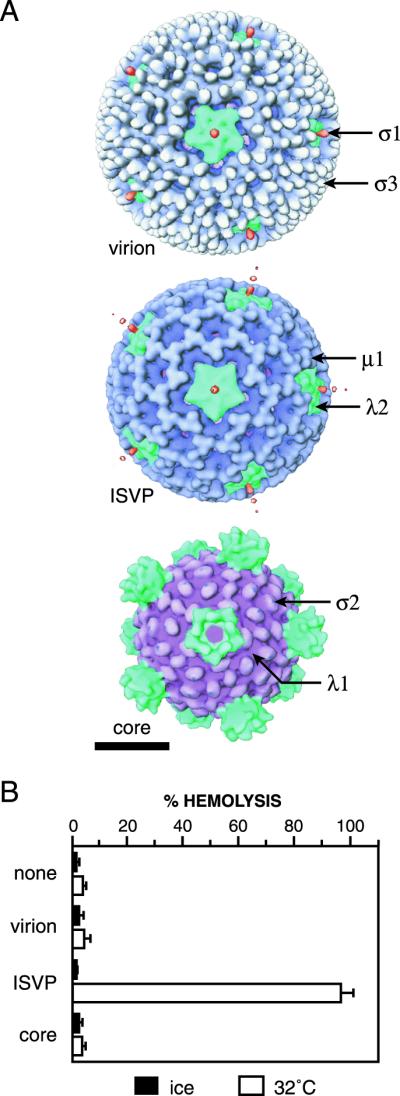
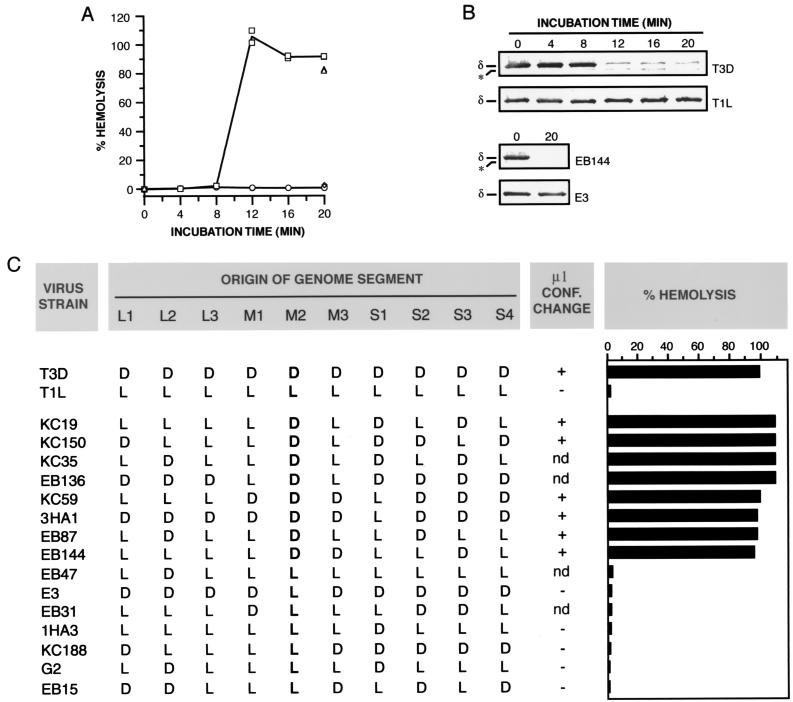
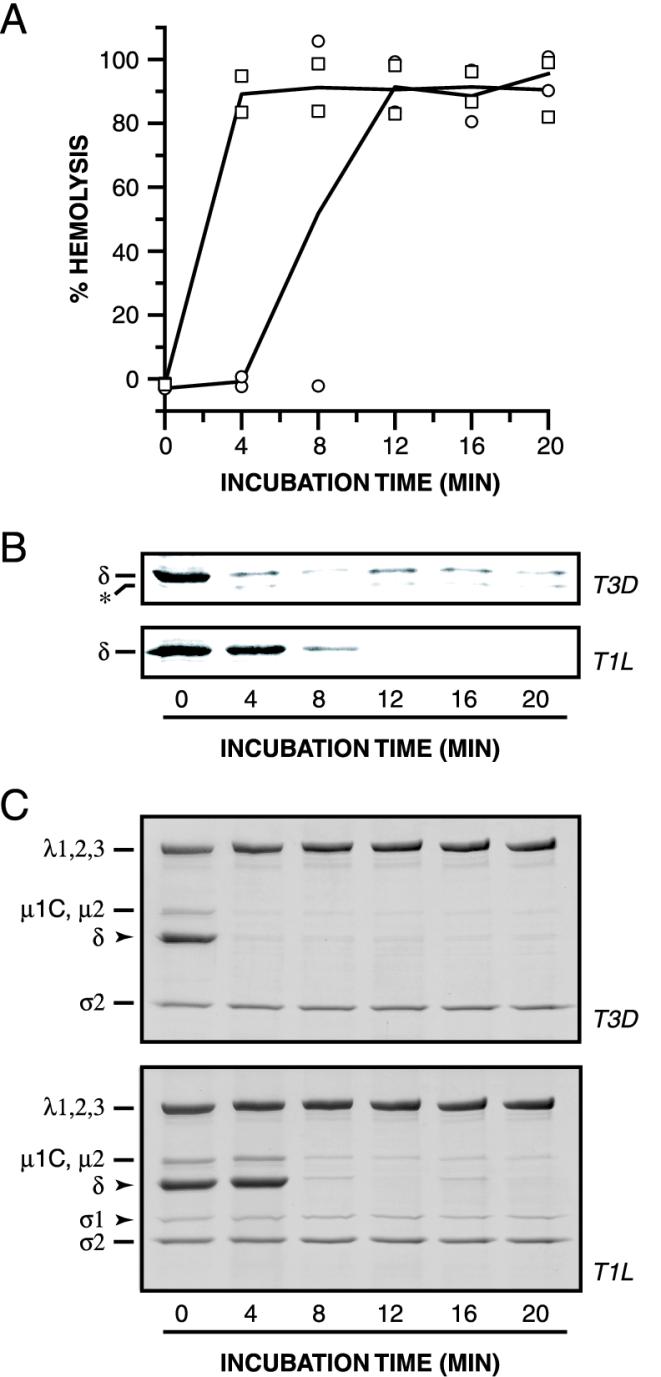
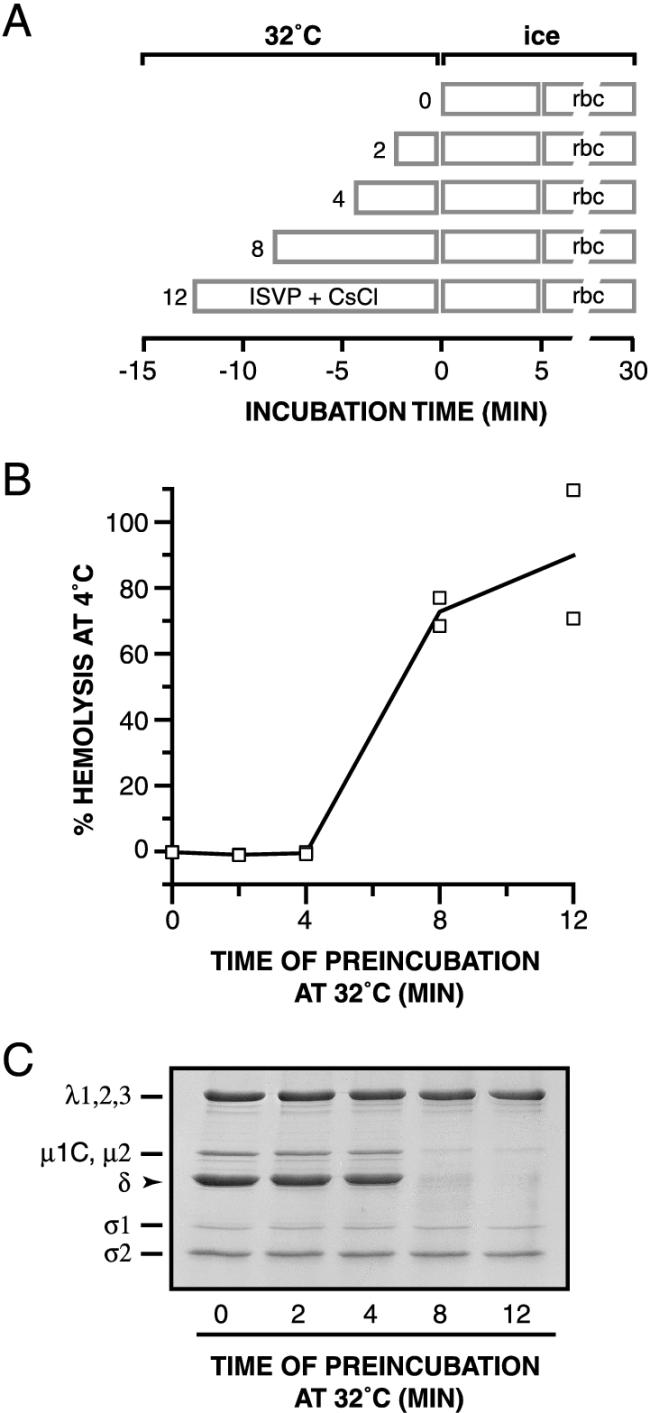
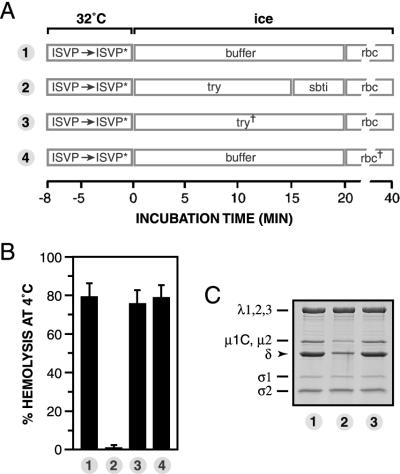
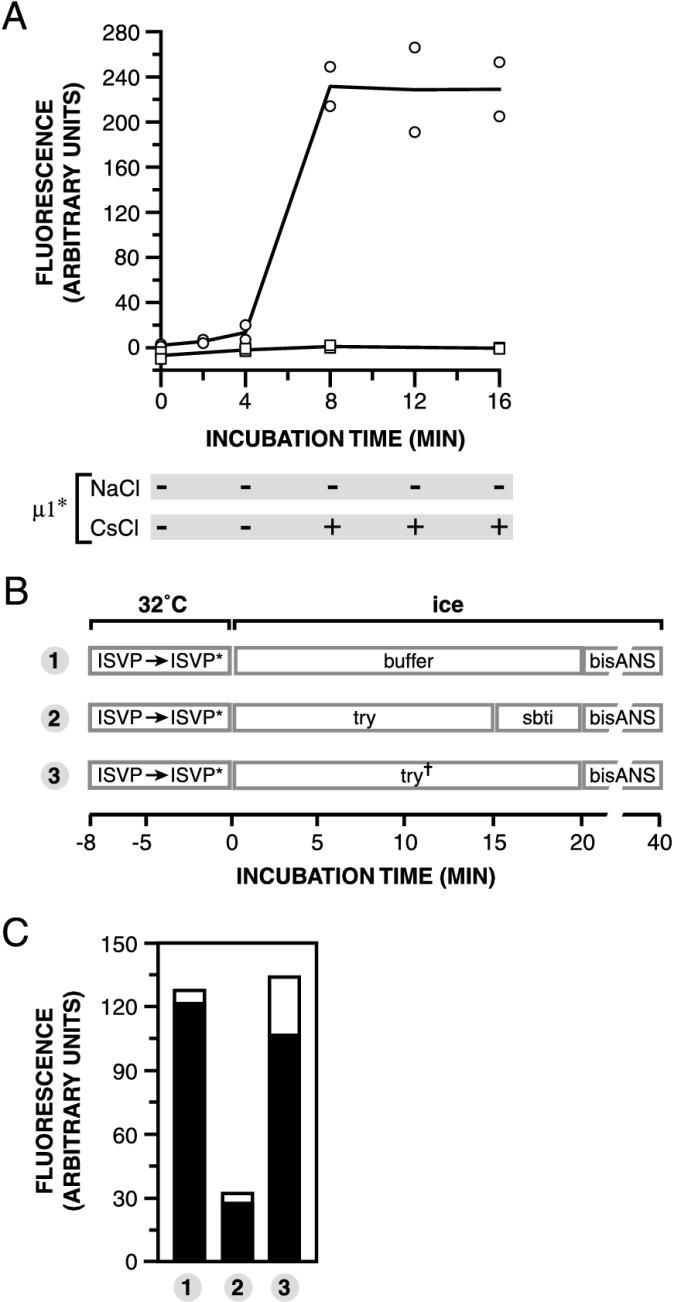
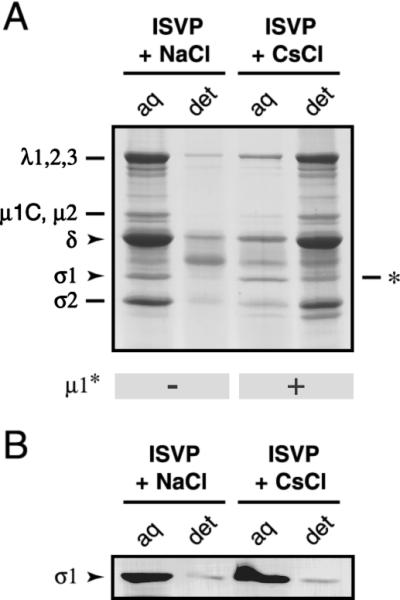
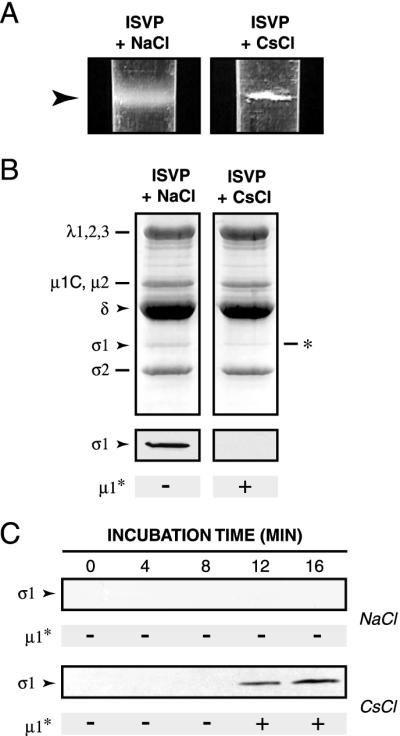
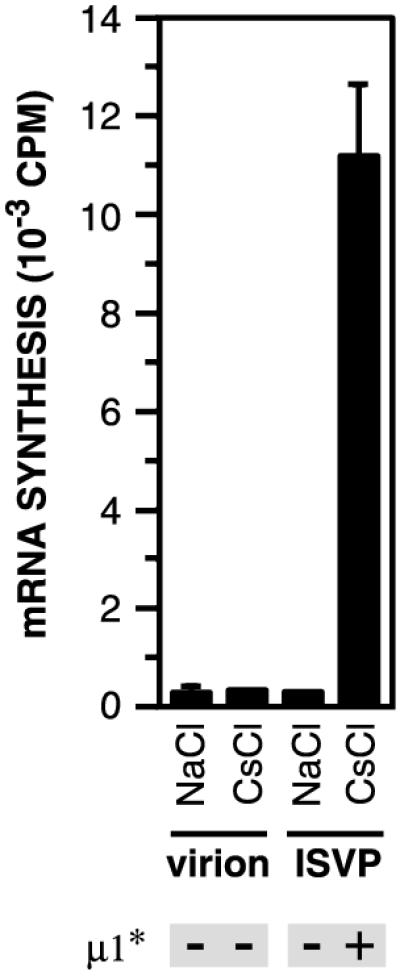
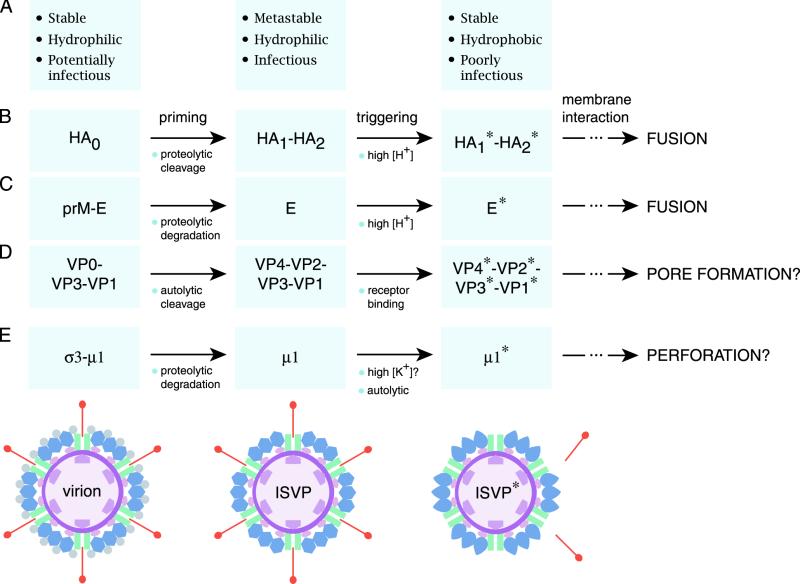
The mechanisms employed by nonenveloped animal viruses to penetrate the membranes of their host cells remain enigmatic. Membrane penetration by the nonenveloped mammalian reoviruses is believed to deliver a partially uncoated, but still large ( approximately 70-nm), particle with active transcriptases for viral mRNA synthesis directly into the cytoplasm. This process is likely initiated by a particle form that resembles infectious subvirion particles (ISVPs), disassembly intermediates produced from virions by proteolytic uncoating. Consistent with that idea, ISVPs, but not virions, can induce disruption of membranes in vitro. Both activities ascribed to ISVP-like particles, membrane disruption in vitro and membrane penetration within cells, are linked to N-myristoylated outer-capsid protein micro 1, present in 600 copies at the surfaces of ISVPs. To understand how micro 1 fulfills its role as the reovirus penetration protein, we monitored changes in ISVPs during the permeabilization of red blood cells induced by these particles. Hemolysis was preceded by a major structural transition in ISVPs, characterized by conformational change in micro 1 and elution of fibrous attachment protein sigma 1. The altered conformer of micro 1 was required for hemolysis and was markedly hydrophobic. The structural transition in ISVPs was further accompanied by derepression of genome-dependent mRNA synthesis by the particle-associated transcriptases. We propose a model for reovirus entry in which (i) primed and triggered conformational changes, analogous to those in enveloped-virus fusion proteins, generate a hydrophobic micro 1 conformer capable of inserting into and disrupting cell membranes and (ii) activation of the viral particles for membrane interaction and mRNA synthesis are concurrent events. Reoviruses provide an opportune system for defining the molecular details of membrane penetration by a large nonenveloped animal virus.
Figures
References
-
- Barton, E. S., J. C. Forrest, J. L. Connolly, J. D. Chappell, Y. Liu, F. J. Schnell, A. Nusrat, C. A. Parkos, and T. S. Dermody. 2001. Junction adhesion molecule is a receptor for reovirus. Cell 104:441-451. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
