

Nitrogen monoxide-mediated control of ferritin synthesis: implications for macrophage iron homeostasis
- PMID: 12209009
- PMCID: PMC129424
- DOI: 10.1073/pnas.192316099
Nitrogen monoxide-mediated control of ferritin synthesis: implications for macrophage iron homeostasis
Abstract
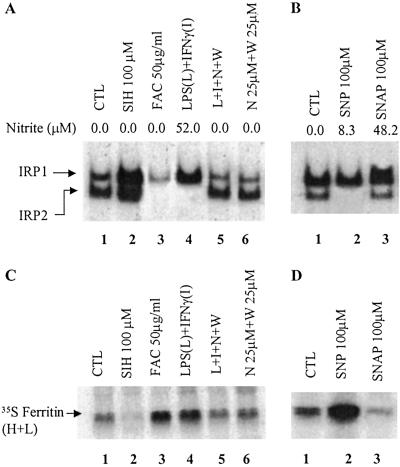
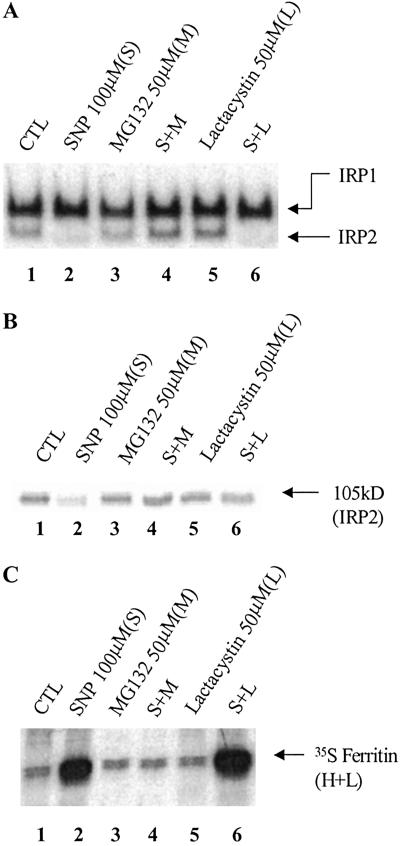
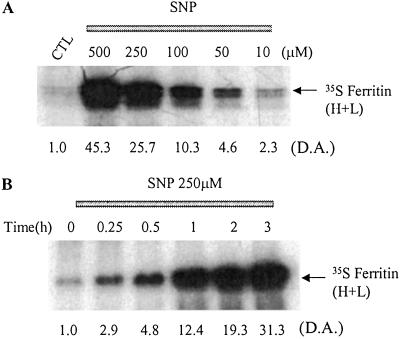
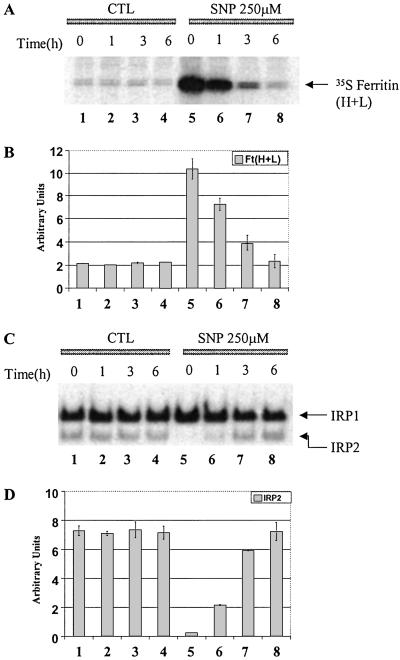
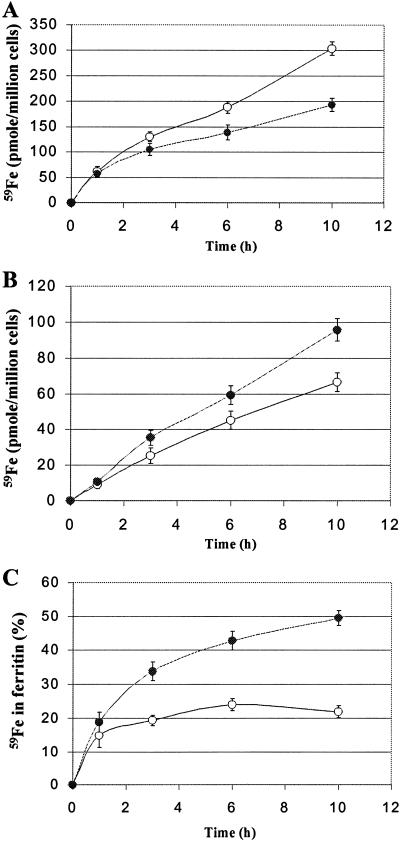
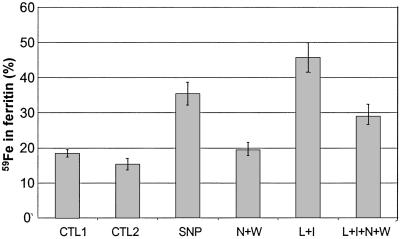
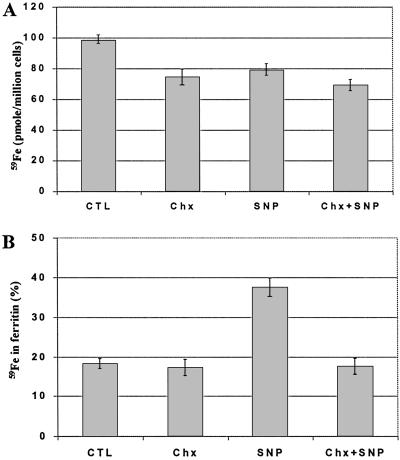
Intracellular iron homeostasis is regulated posttranscriptionally by iron regulatory proteins 1 and 2 (IRP1 and IRP2). In the absence of iron in the labile pool, IRPs bind to specific nucleotide sequences called iron responsive elements (IREs), which are located in the 5' untranslated region of ferritin mRNA and the 3' untranslated region of transferrin receptor mRNA. IRP binding to the IREs suppresses ferritin translation and stabilizes transferrin receptor mRNA, whereas the opposite scenario develops in iron-replete cells. Binding of IRPs to the IREs is also affected by nitrogen monoxide (NO), but there are conflicting reports regarding the effect of NO on ferritin synthesis. In this study, we demonstrated that a short exposure of RAW 264.7 cells (a macrophage cell line) to the NO+ donor, sodium nitroprusside (SNP), resulted in a dramatic increase in ferritin synthesis. The SNP-mediated increase of ferritin synthesis could be blocked by MG132, an inhibitor of proteasome-dependent protein degradation, which also prevented the degradation of IRP2 caused by SNP treatment. Moreover, treatment of RAW 264.7 cells with IFN-gamma and lipopolysaccharide caused IRP2 degradation and stimulated ferritin synthesis, changes that could be prevented by specific inhibitors of inducible nitric oxide synthase. Furthermore, the SNP-mediated increase in ferritin synthesis was associated with a significant enhancement of iron incorporation into ferritin. These observations indicate that NO+-mediated modulation of IRP2 plays an important role in controlling ferritin synthesis and iron metabolism in murine macrophages.
Figures
Similar articles
-
Iron regulatory protein-independent regulation of ferritin synthesis by nitrogen monoxide.FEBS J. 2006 Aug;273(16):3828-36. doi: 10.1111/j.1742-4658.2006.05390.x. FEBS J. 2006. PMID: 16911529
-
Effects of interferon-gamma and lipopolysaccharide on macrophage iron metabolism are mediated by nitric oxide-induced degradation of iron regulatory protein 2.J Biol Chem. 2000 Mar 3;275(9):6220-6. doi: 10.1074/jbc.275.9.6220. J Biol Chem. 2000. PMID: 10692416
-
Control of transferrin receptor expression via nitric oxide-mediated modulation of iron-regulatory protein 2.J Biol Chem. 1999 Nov 12;274(46):33035-42. doi: 10.1074/jbc.274.46.33035. J Biol Chem. 1999. PMID: 10551872
-
Nitric oxide-mediated modulation of iron regulatory proteins: implication for cellular iron homeostasis.Blood Cells Mol Dis. 2002 Nov-Dec;29(3):400-10. doi: 10.1006/bcmd.2002.0579. Blood Cells Mol Dis. 2002. PMID: 12547230 Review.
-
Role of nitric oxide in cellular iron metabolism.Biometals. 2003 Mar;16(1):125-35. doi: 10.1023/a:1020788603046. Biometals. 2003. PMID: 12572672 Review.
Cited by
-
Lysosomal iron modulates NMDA receptor-mediated excitation via small GTPase, Dexras1.Mol Brain. 2016 Apr 14;9:38. doi: 10.1186/s13041-016-0220-8. Mol Brain. 2016. PMID: 27080392 Free PMC article.
-
Tetrandrine ameliorates cirrhosis and portal hypertension by inhibiting nitric oxide in cirrhotic rats.J Huazhong Univ Sci Technolog Med Sci. 2004;24(4):385-8, 395. doi: 10.1007/BF02861874. J Huazhong Univ Sci Technolog Med Sci. 2004. PMID: 15587405
-
Inflaming the Brain with Iron.Antioxidants (Basel). 2021 Jan 6;10(1):61. doi: 10.3390/antiox10010061. Antioxidants (Basel). 2021. PMID: 33419006 Free PMC article. Review.
-
Aberrant Transferrin and Ferritin Upregulation Elicits Iron Accumulation and Oxidative Inflammaging Causing Ferroptosis and Undermines Estradiol Biosynthesis in Aging Rat Ovaries by Upregulating NF-Κb-Activated Inducible Nitric Oxide Synthase: First Demonstration of an Intricate Mechanism.Int J Mol Sci. 2022 Oct 21;23(20):12689. doi: 10.3390/ijms232012689. Int J Mol Sci. 2022. PMID: 36293552 Free PMC article.
-
Iron regulatory protein 1 outcompetes iron regulatory protein 2 in regulating cellular iron homeostasis in response to nitric oxide.J Biol Chem. 2011 Jul 1;286(26):22846-54. doi: 10.1074/jbc.M111.231902. Epub 2011 May 12. J Biol Chem. 2011. PMID: 21566147 Free PMC article.
References
-
- Munro H. N. & Linder, M. C. (1978) Physiol. Rev. 58, 317-396. - PubMed
-
- Harrison P. M. & Arosio, P. (1996) Biochim. Biophys. Acta 1275, 161-203. - PubMed
-
- Levi S., Luzzago, A., Cozzi, A., Franceschinelli, F., Albertini, A. & Arosio, P. (1988) J. Biol. Chem. 263, 18086-18092. - PubMed
-
- Santambrogio P., Levi, S., Arosio, P., Palagi, L., Vecchio, G., Lawson, D. M., Yewdall, S. J., Artymium, P. J., Harrison, P. M., Jappelli, R. & Cesareni, G. (1992) J. Biol. Chem. 267, 14077-14083. - PubMed
-
- Levi S., Santambrogio, P., Cozzi, A., Rovida, E., Corsi, B., Tamborini, E., Spada, S., Albertini, A. & Arosio, P. (1994) J. Mol. Biol. 238, 649-654. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
