

Position of aromatic residues in the S6 domain, not inactivation, dictates cisapride sensitivity of HERG and eag potassium channels
- PMID: 12209010
- PMCID: PMC129467
- DOI: 10.1073/pnas.192367299
Position of aromatic residues in the S6 domain, not inactivation, dictates cisapride sensitivity of HERG and eag potassium channels
Abstract
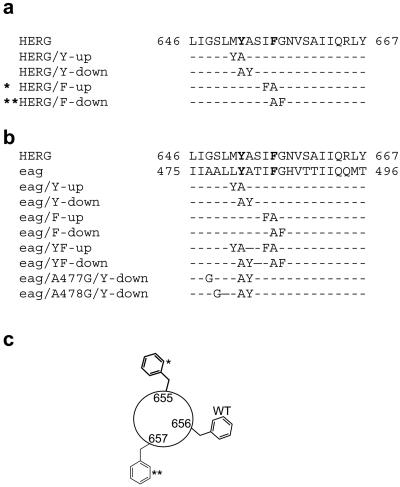
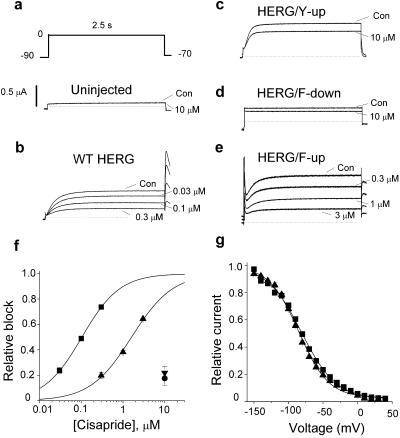
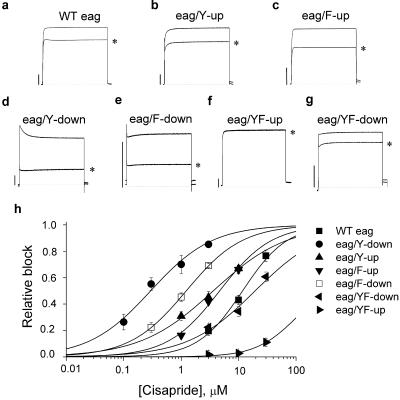
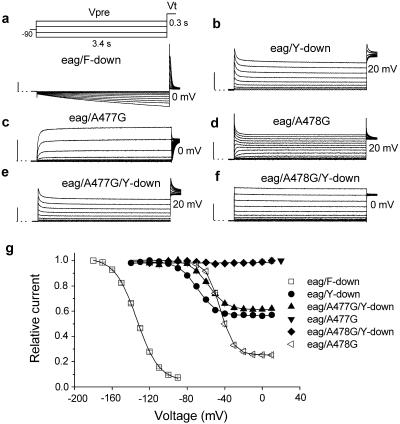
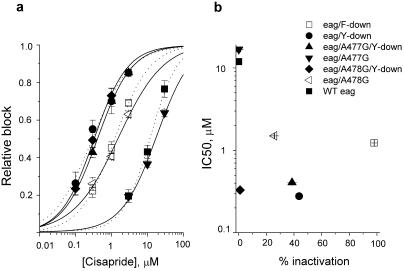
Unintended block of HERG K+ channels is a side effect of many common medications and is the most common cause of acquired long QT syndrome associated with increased risk of life-threatening arrhythmias. The molecular mechanism of high-affinity HERG block by structurally diverse compounds has been attributed to pi-stacking and cation-pi interactions of a drug (e.g., cisapride) with specific aromatic amino acid residues (Tyr-652 and Phe-656) in the S6 alpha-helical domain that face the central cavity of the channel. It also has been proposed that strong C-type inactivation of HERG facilitates or is the primary determinant of high-affinity drug binding. The structurally related, but noninactivating eag channel is insensitive to HERG blockers unless inactivation is induced by specific amino acid mutations [Ficker, E., Jarolimek, W. & Brown, A. M. (2001) Mol. Pharmacol. 60, 1343-1348]. Here we examine the relative importance of inactivation vs. positioning of S6 aromatic residues in determining sensitivity of HERG and eag channels to block by cisapride. The repositioning of Tyr-652 or Phe-656 along the S6 alpha-helical domain of HERG reduced sensitivity of channels to block by cisapride. Moreover, independent of inactivation, repositioning of the equivalent aromatic residues in Drosophila eag channels induced sensitivity to block by cisapride. These findings suggest that positioning of S6 aromatic residues relative to the central cavity of the channel, not inactivation per se determines drug block of HERG or eag channels.
Figures
References
-
- Sanguinetti M. C., Jiang, C., Curran, M. E. & Keating, M. T. (1995) Cell 81, 299-307. - PubMed
-
- Trudeau M., Warmke, J. W., Ganetzky, B. & Robertson, G. A. (1995) Science 269, 92-95. - PubMed
-
- Curran M. E., Splawski, I., Timothy, K. W., Vincent, G. M., Green, E. D. & Keating, M. T. (1995) Cell 80, 795-803. - PubMed
-
- Roden D. M., Lazzara, R., Rosen, M., Schwartz, P. J., Towbin, J. & Vincent, G. M. (1996) Circulation 94, 1996-2012. - PubMed
-
- Keating M. T. & Sanguinetti, M. C. (2001) Cell 104, 569-580. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
