

Previously uncharacterized isoforms of divalent metal transporter (DMT)-1: implications for regulation and cellular function
- PMID: 12209011
- PMCID: PMC129447
- DOI: 10.1073/pnas.192423399
Previously uncharacterized isoforms of divalent metal transporter (DMT)-1: implications for regulation and cellular function
Abstract
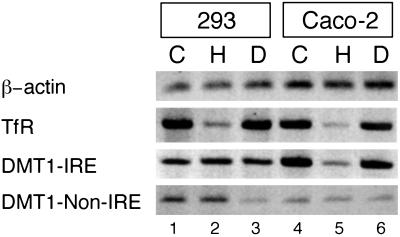
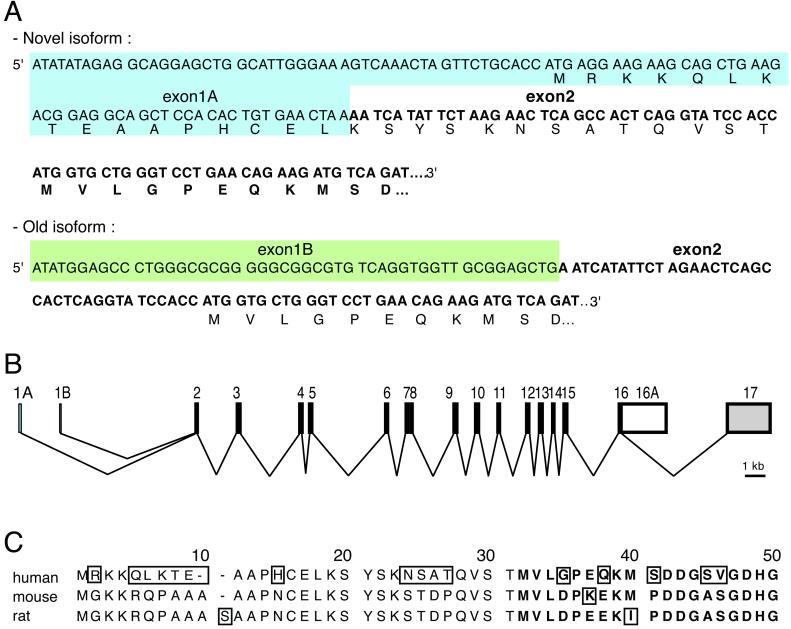
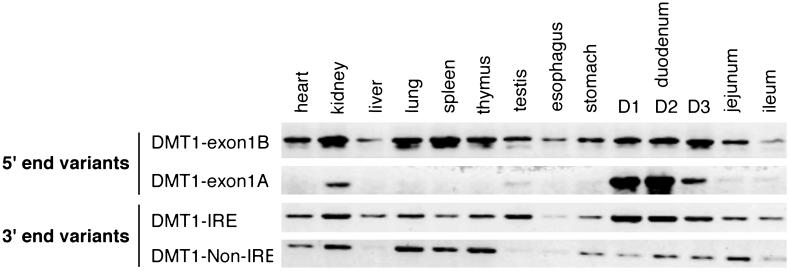
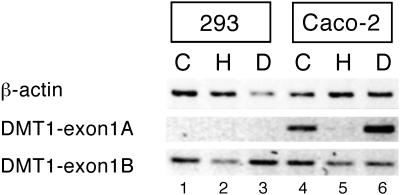
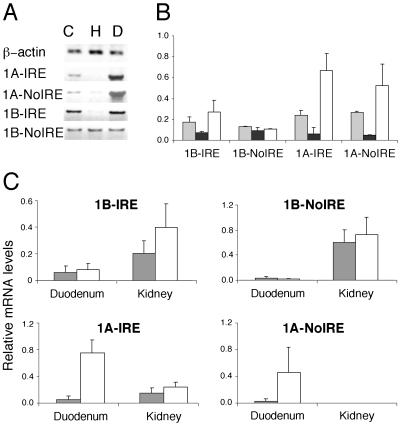
Divalent metal transporter 1 (DMT1) mediates apical iron uptake into duodenal enterocytes and also transfers iron from the endosome into the cytosol after cellular uptake via the transferrin receptor. Hence, mutations in DMT1 cause systemic iron deficiency and anemia. DMT1 mRNA levels are increased in the duodenum of iron-deficient animals. This regulation has been observed for DMT1 mRNA harboring an iron-responsive element (IRE) in its 3' UTR, but not for a processing variant lacking a 3'UTR IRE, suggesting that the IRE regulates the expression of DMT1 mRNA in response to iron levels. Here, we show that iron regulation of DMT1 involves the expression of a previously unrecognized upstream 5' exon (exon 1A) of the human and murine DMT1 gene. The expression of this previously uncharacterized 5' exon is tissue-specific and particularly prevalent in the duodenum and kidney. It adds an in-frame AUG translation initiation codon extending the DMT1 ORF by a conserved sequence of 29-31 amino acids. In combination with the IRE- and non-IRE variants in the 3'UTR, our results reveal the existence of four DMT1 mRNA isoforms predicting the synthesis of four different DMT1 proteins. We show that two regulatory regions, the 5' promoter/exon 1A region and the IRE-containing terminal exon participate in iron regulation of DMT1 expression, which operate in a tissue-specific way. These results uncover an unexpected complexity of DMT1 expression and regulation, with implications for understanding the physiology, cell biology, and pathophysiology of mammalian iron metabolism.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
