

Reproductive pathology and sperm physiology in acid sphingomyelinase-deficient mice
- PMID: 12213735
- PMCID: PMC1867239
- DOI: 10.1016/S0002-9440(10)64267-8
Reproductive pathology and sperm physiology in acid sphingomyelinase-deficient mice
Abstract
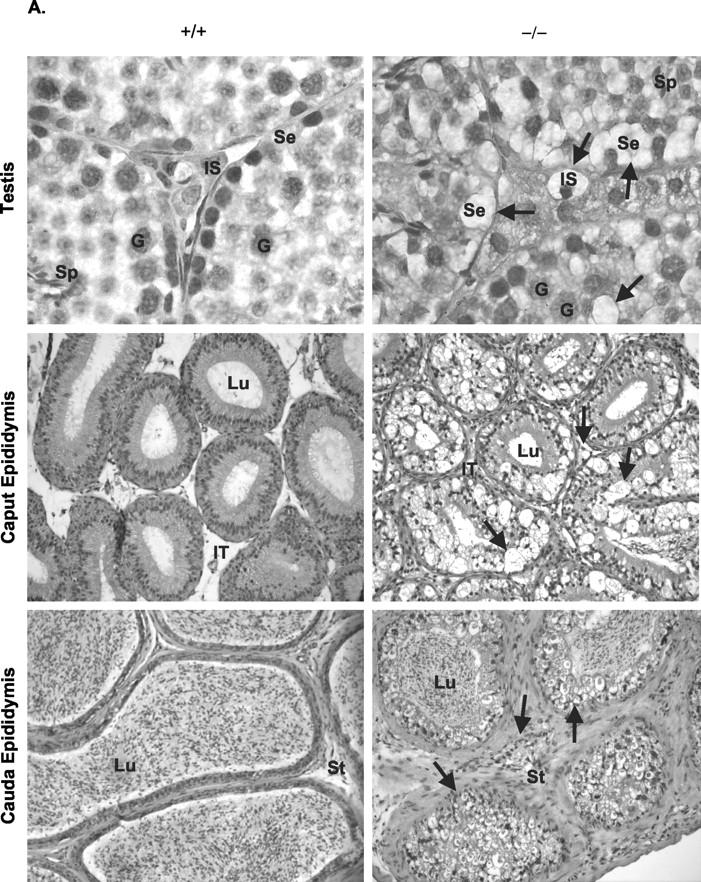
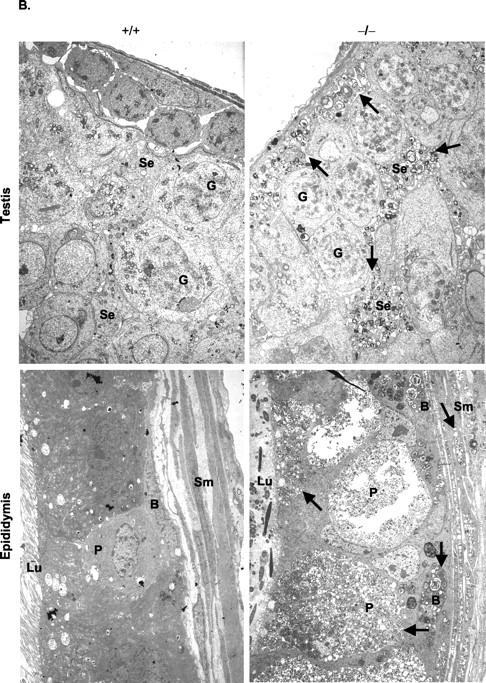
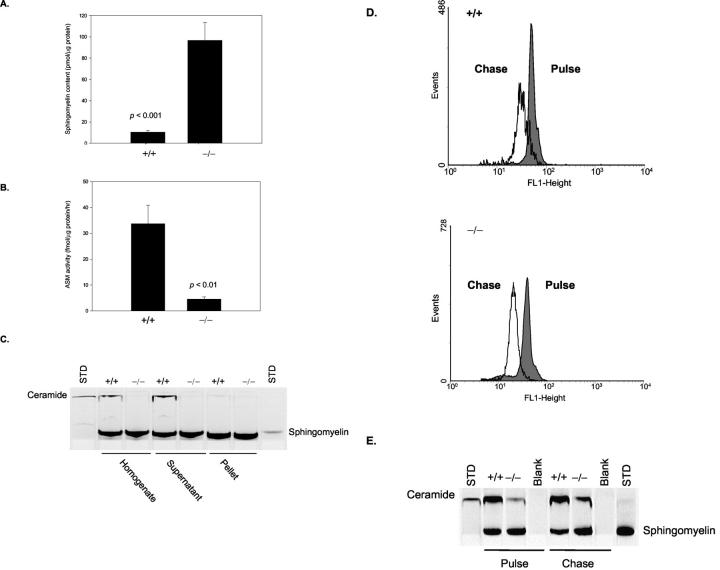
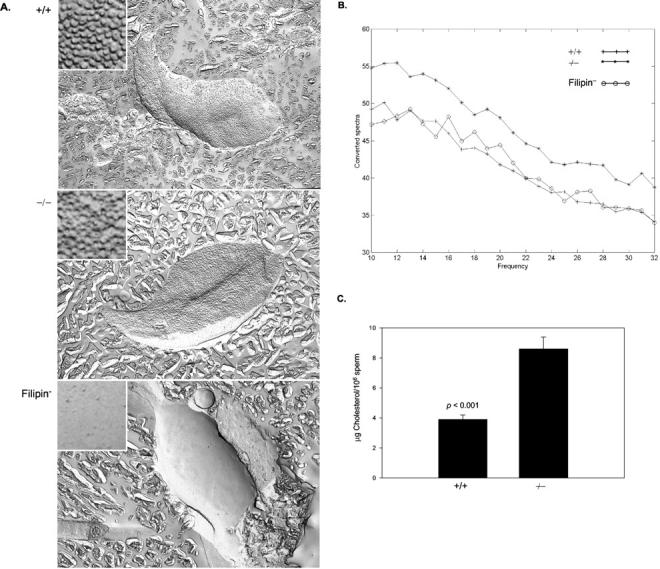
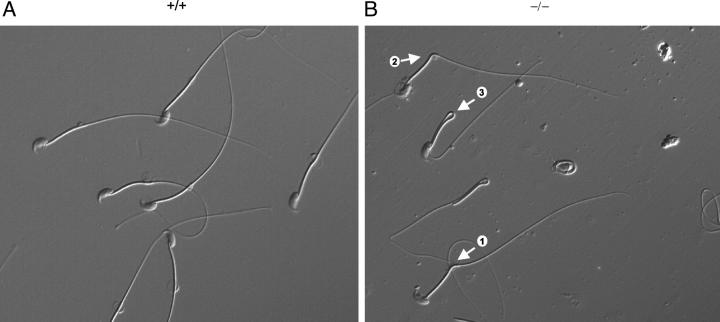
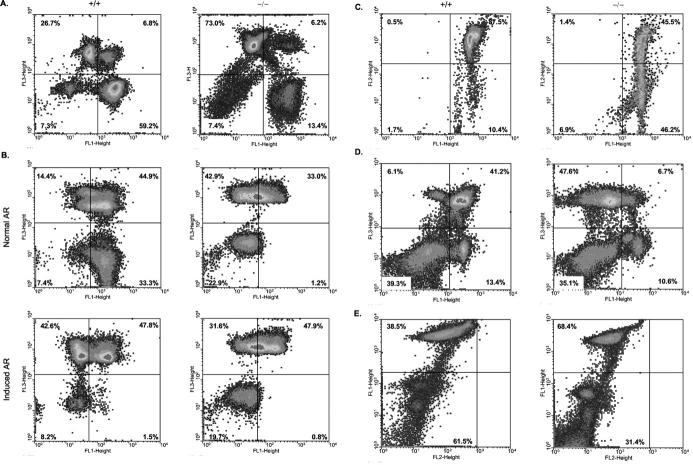
Types A and B Niemann-Pick disease (NPD) are lysosomal storage disorders resulting from the deficient activity of acid sphingomyelinase (ASM). In this manuscript we report the pathobiology of male gonadal tissue and sperm in a knockout mouse model of NPD and demonstrate the importance of ASM for normal sperm maturation and function. Characteristic lipid-filled vacuoles were evident in light micrographs of testis' seminiferous tubules and epithelial cells lining the epididymis of -/- mice. Electron micrographs extended these findings and revealed storage vesicles within Sertoli cells of the seminiferous tubules. Mature spermatozoa from -/- mice showed marked ASM deficiency and elevated levels of sphingomyelin and cholesterol. Flow cytometric analysis revealed that affected spermatozoa had disrupted plasma and acrosome membranes, and mitochondrial membrane depolarization. They also did not undergo proper capacitation. Morphological abnormalities such as kinks and bends at the midpiece-principle piece junction were evident in spermatozoa from affected mice, with consequent deficits in motility. Notably, the mutant sperm regained normal morphology on incubation in mild detergent, demonstrating that the bending defects were a direct consequence of membrane lipid accumulation. A mechanism for these abnormalities is proposed that suggests lipid accumulation in the gonads results in regulatory volume decrease defects within the developing sperm, and that regulatory volume decrease defects, in turn, lead to the observed abnormalities in sperm morphology and function. These results provide in vivo evidence that ASM activity plays a critical role in sperm maturation and function, and a basis for similar studies in sexually mature, male NPD patients.
Figures
References
-
- Schuchman E, Desnick R: Niemann-Pick disease types A and B: acid sphingomyelinase deficiencies. Scriver C Beaudet A Sly W Valle D eds. The Metabolic and Molecular Basis of Inherited Disease, ed 8 2001:p 3589 McGraw-Hill, New York
-
- Levade T, Jaffrezou JP: Signaling sphingomyelinases: which, where, how and why? Biochim Biophys Acta 1999, 1438:1-17 - PubMed
-
- Tomiuk S, Zumbansen M, Stoffel W: Characterization and subcellular localization of murine and human magnesium-dependent neutral sphingomyelinase. J Biol Chem 2000, 275:5710-5717 - PubMed
-
- Schuchman EH, Suchi M, Takahashi T, Sandoff K, Desnick RJ: Human acid sphingomyelinase. Isolation, nucleotide sequence, and expression of the full-length and alternatively spliced cDNAs. J Biol Chem 1991, 13:8531-8539 - PubMed
-
- Schuchman EH, Levran O, Pereira LV, Desnick RJ: Structural organization and complete nucleotide sequence of the gene encoding human acid sphingomyelinase (SMPD1). Genomics 1992, 12:197-205 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
