

Regulation of membrane fusion by the membrane-proximal coil of the t-SNARE during zippering of SNAREpins
- PMID: 12213837
- PMCID: PMC2173141
- DOI: 10.1083/jcb.200112081
Regulation of membrane fusion by the membrane-proximal coil of the t-SNARE during zippering of SNAREpins
Abstract
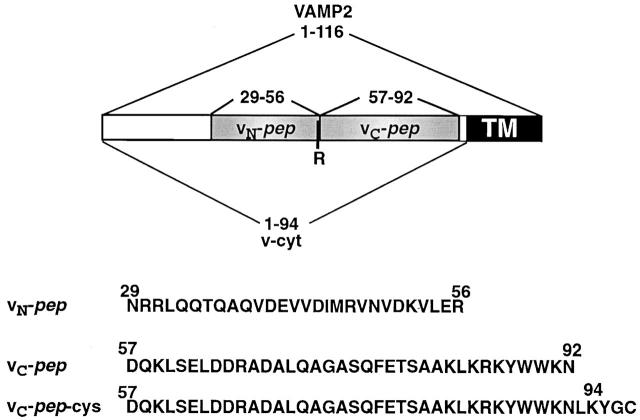
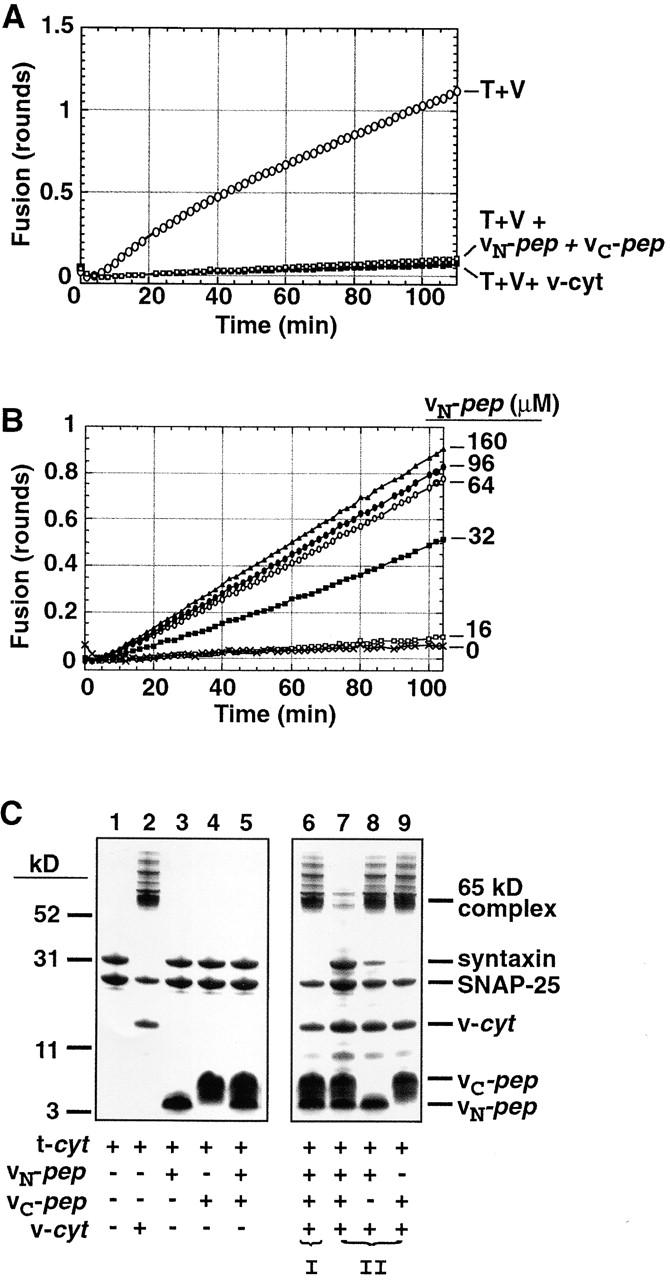
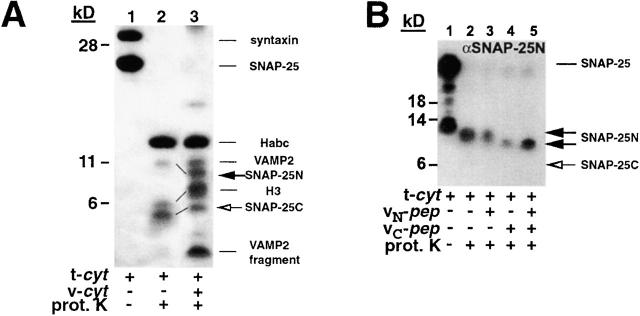
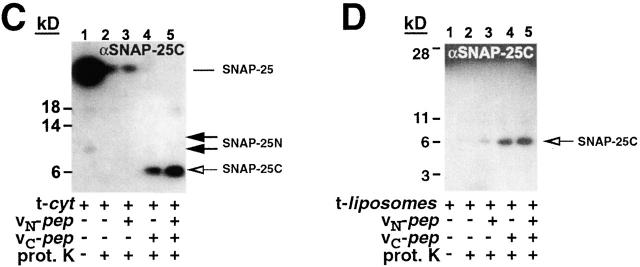
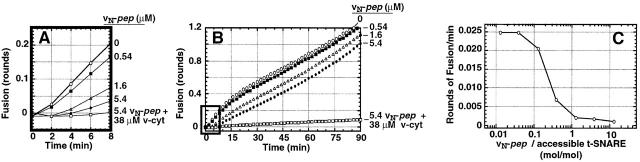
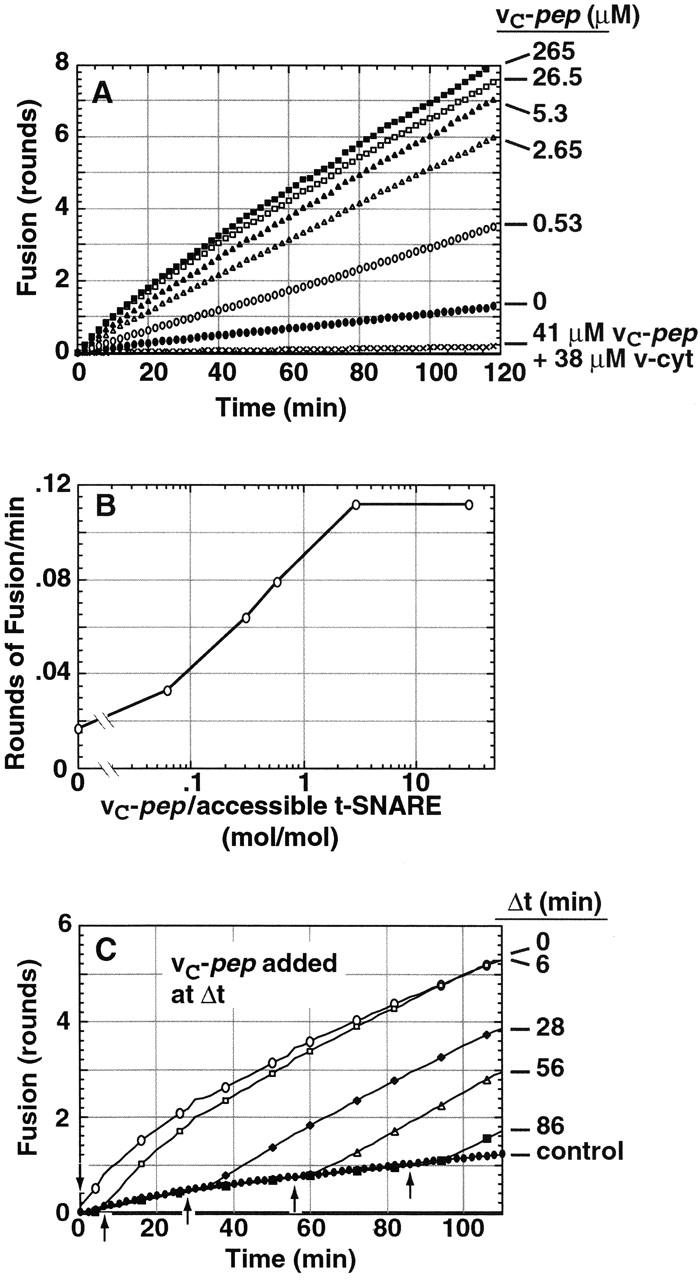
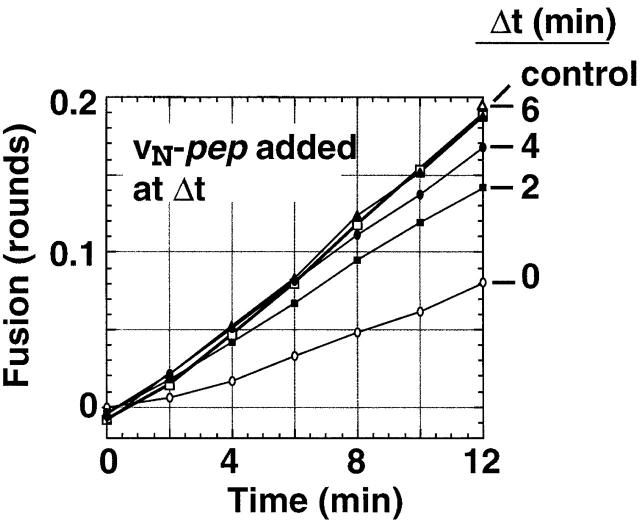
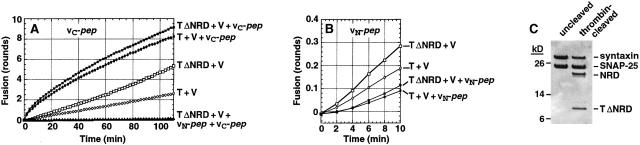
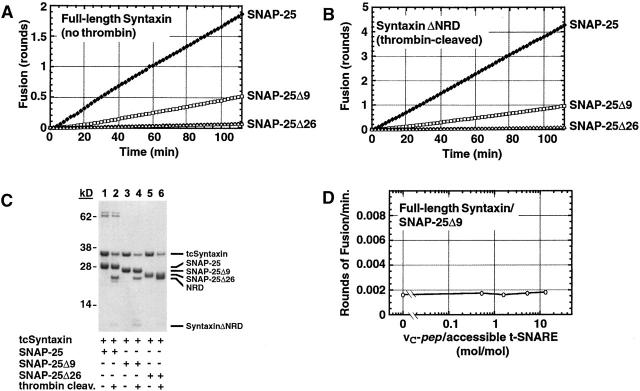
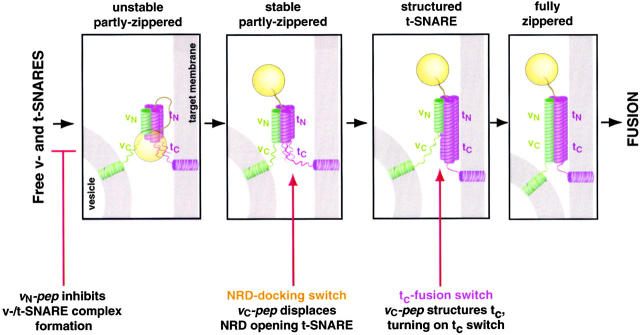
We utilize structurally targeted peptides to identify a "tC fusion switch" inherent to the coil domains of the neuronal t-SNARE that pairs with the cognate v-SNARE. The tC fusion switch is located in the membrane-proximal portion of the t-SNARE and controls the rate at which the helical bundle that forms the SNAREpin can zip up to drive bilayer fusion. When the fusion switch is "off" (the intrinsic state of the t-SNARE), zippering of the helices from their membrane-distal ends is impeded and fusion is slow. When the tC fusion switch is "on," fusion is much faster. The tC fusion switch can be thrown by a peptide that corresponds to the membrane-proximal half of the cognate v-SNARE, and binds reversibly to the cognate region of the t-SNARE. This structures the coil in the membrane-proximal domain of the t-SNARE and accelerates fusion, implying that the intrinsically unstable coil in that region is a natural impediment to the completion of zippering, and thus, fusion. Proteins that stabilize or destabilize one or the other state of the tC fusion switch would exert fine temporal control over the rate of fusion after SNAREs have already partly zippered up.
Figures
References
-
- Betz, A., M. Okamoto, F. Benseler, and N. Brose. 1997. Direct interaction of the rat unc-13 homologue Munc 13-1 with the N-terminus of syntaxin. J. Biol. Chem. 272:2520–2526. - PubMed
-
- Chan, D.C., and P.S. Kim. 1998. HIV entry and its inhibition. Cell. 93:681–684. - PubMed
-
- Chen, X., D.R. Tomchick, E. Kovrigin, D. Arac, M. Machius, T.C. Sudhof, and J. Rizo. 2002. Three-dimensional structure of the complexin/SNARE complex. Neuron. 33:397–409. - PubMed
-
- Chen, Y.A., S.J. Scales, S.M. Patel, Y.-C. Doung, and R.H. Scheller. 1999. SNARE complex formation is triggered by Ca2+ and drives membrane fusion. Cell. 97:165–174. - PubMed
