

KOBITO1 encodes a novel plasma membrane protein necessary for normal synthesis of cellulose during cell expansion in Arabidopsis
- PMID: 12215501
- PMCID: PMC150751
- DOI: 10.1105/tpc.002873
KOBITO1 encodes a novel plasma membrane protein necessary for normal synthesis of cellulose during cell expansion in Arabidopsis
Abstract
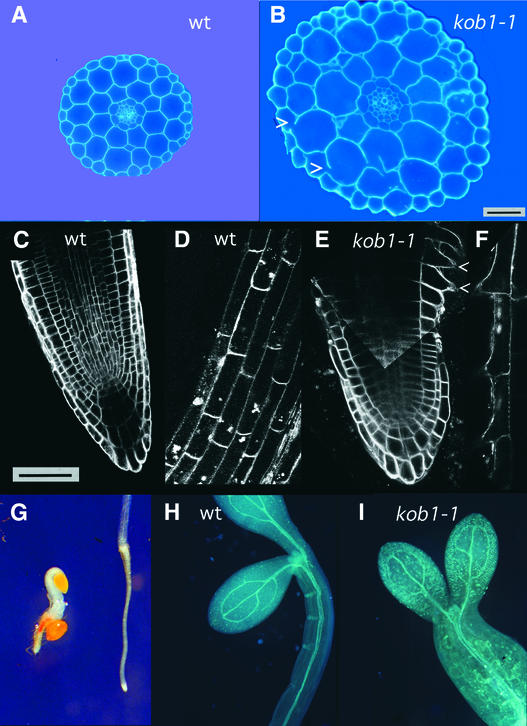
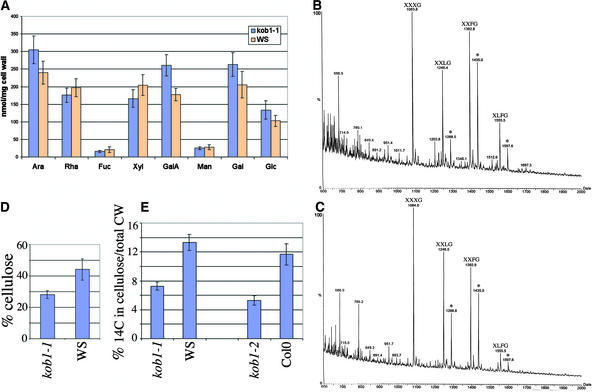
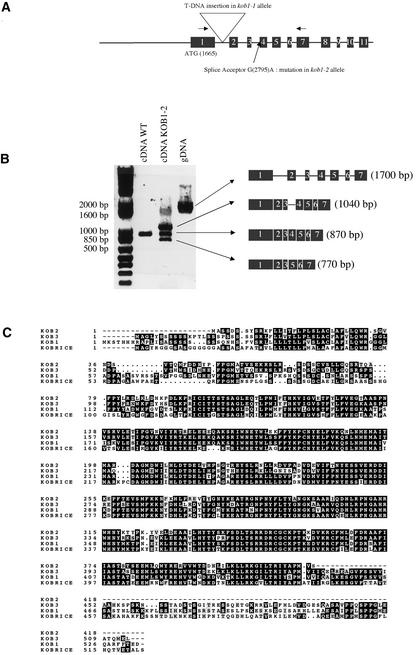
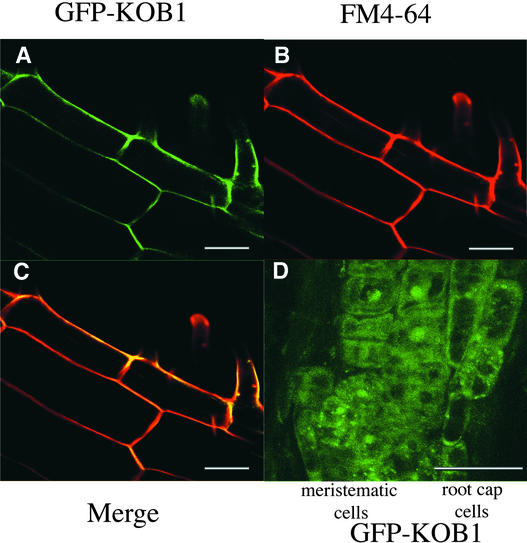
The cell wall is the major limiting factor for plant growth. Wall extension is thought to result from the loosening of its structure. However, it is not known how this is coordinated with wall synthesis. We have identified two novel allelic cellulose-deficient dwarf mutants, kobito1-1 and kobito1-2 (kob1-1 and kob1-2). The cellulose deficiency was confirmed by the direct observation of microfibrils in most recent wall layers of elongating root cells. In contrast to the wild type, which showed transversely oriented parallel microfibrils, kob1 microfibrils were randomized and occluded by a layer of pectic material. No such changes were observed in another dwarf mutant, pom1, suggesting that the cellulose defect in kob1 is not an indirect result of the reduced cell elongation. Interestingly, in the meristematic zone of kob1 roots, microfibrils appeared unaltered compared with the wild type, suggesting a role for KOB1 preferentially in rapidly elongating cells. KOB1 was cloned and encodes a novel, highly conserved, plant-specific protein that is plasma membrane bound, as shown with a green fluorescent protein-KOB1 fusion protein. KOB1 mRNA was present in all organs investigated, and its overexpression did not cause visible phenotypic changes. KOB1 may be part of the cellulose synthesis machinery in elongating cells, or it may play a role in the coordination between cell elongation and cellulose synthesis.
Figures


Similar articles
-
The elongation defective1 mutant of Arabidopsis is impaired in the gene encoding a serine-rich secreted protein.Plant Mol Biol. 2003 Nov;53(4):581-95. doi: 10.1023/B:PLAN.0000019067.05185.d6. Plant Mol Biol. 2003. PMID: 15010620
-
Glycosyltransferase-like protein ABI8/ELD1/KOB1 promotes Arabidopsis hypocotyl elongation through regulating cellulose biosynthesis.Plant Cell Environ. 2015 Mar;38(3):411-22. doi: 10.1111/pce.12395. Epub 2014 Aug 5. Plant Cell Environ. 2015. PMID: 24995569
-
Regulation of plasmodesmatal permeability and stomatal patterning by the glycosyltransferase-like protein KOBITO1.Plant Physiol. 2012 May;159(1):156-68. doi: 10.1104/pp.112.194563. Epub 2012 Mar 28. Plant Physiol. 2012. PMID: 22457425 Free PMC article.
-
Cellulose microfibrils in plants: biosynthesis, deposition, and integration into the cell wall.Int Rev Cytol. 2000;199:161-99. doi: 10.1016/s0074-7696(00)99004-1. Int Rev Cytol. 2000. PMID: 10874579 Review.
-
Biosynthesis and properties of the plant cell wall.Curr Opin Plant Biol. 2002 Dec;5(6):536-42. doi: 10.1016/s1369-5266(02)00306-0. Curr Opin Plant Biol. 2002. PMID: 12393017 Review.
Cited by
-
Plasmodesmata dynamics are coordinated by intracellular signaling pathways.Curr Opin Plant Biol. 2013 Oct;16(5):614-20. doi: 10.1016/j.pbi.2013.07.007. Epub 2013 Aug 23. Curr Opin Plant Biol. 2013. PMID: 23978390 Free PMC article. Review.
-
Callose homeostasis at plasmodesmata: molecular regulators and developmental relevance.Front Plant Sci. 2014 Apr 21;5:138. doi: 10.3389/fpls.2014.00138. eCollection 2014. Front Plant Sci. 2014. PMID: 24795733 Free PMC article.
-
Genome-wide analysis of the cellulose toolbox of Primulina eburnea, a calcium-rich vegetable.BMC Plant Biol. 2023 May 16;23(1):259. doi: 10.1186/s12870-023-04266-z. BMC Plant Biol. 2023. PMID: 37189063 Free PMC article.
-
Abscisic Acid: Hidden Architect of Root System Structure.Plants (Basel). 2015 Aug 11;4(3):548-72. doi: 10.3390/plants4030548. Plants (Basel). 2015. PMID: 27135341 Free PMC article. Review.
-
Combined full-length transcriptomic and metabolomic analysis reveals the regulatory mechanisms of adaptation to salt stress in asparagus.Front Plant Sci. 2022 Oct 27;13:1050840. doi: 10.3389/fpls.2022.1050840. eCollection 2022. Front Plant Sci. 2022. PMID: 36388563 Free PMC article.
References
-
- Arioli, T., et al. (1998). Molecular analysis of cellulose biosynthesis in Arabidopsis. Science 279, 717–720. - PubMed
-
- Brummell, D.A., and Hall, J.L. (1985). The role of cell wall synthesis in sustained auxin-induced growth. Physiol. Plant. 63, 406–412.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
