

Site-directed mutagenesis of conserved charged amino acid residues in ClpB from Escherichia coli
- PMID: 12220194
- PMCID: PMC1851691
- DOI: 10.1021/bi026161s
Site-directed mutagenesis of conserved charged amino acid residues in ClpB from Escherichia coli
Abstract
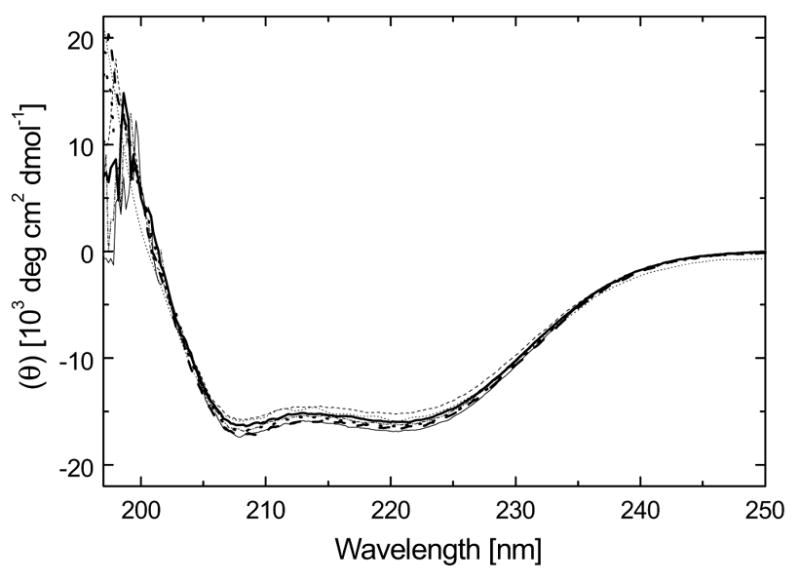
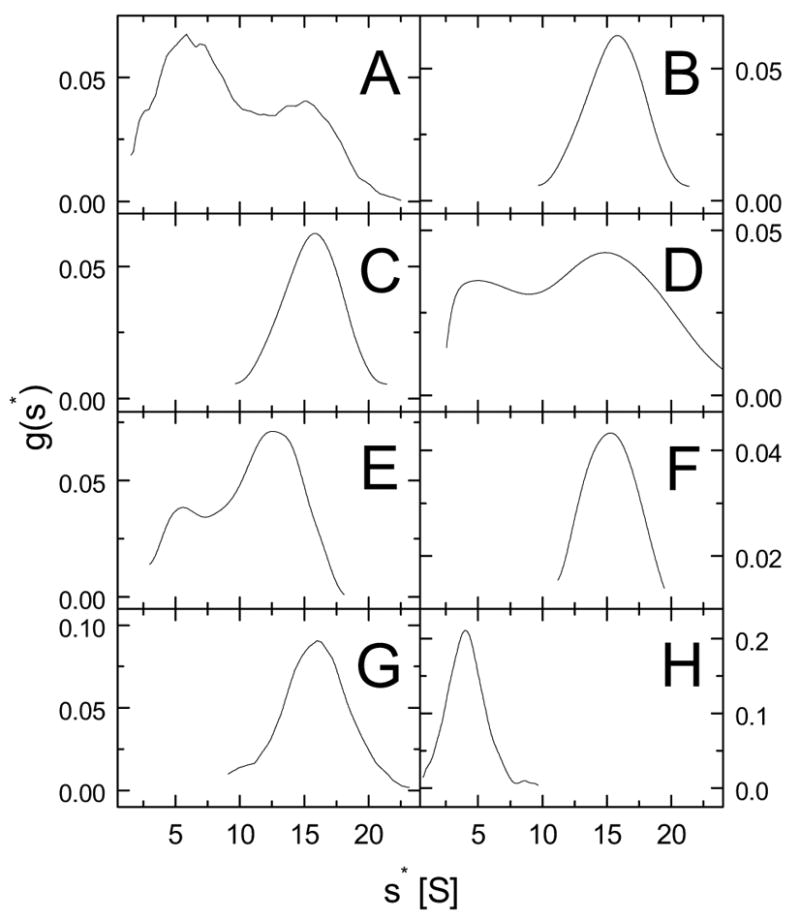
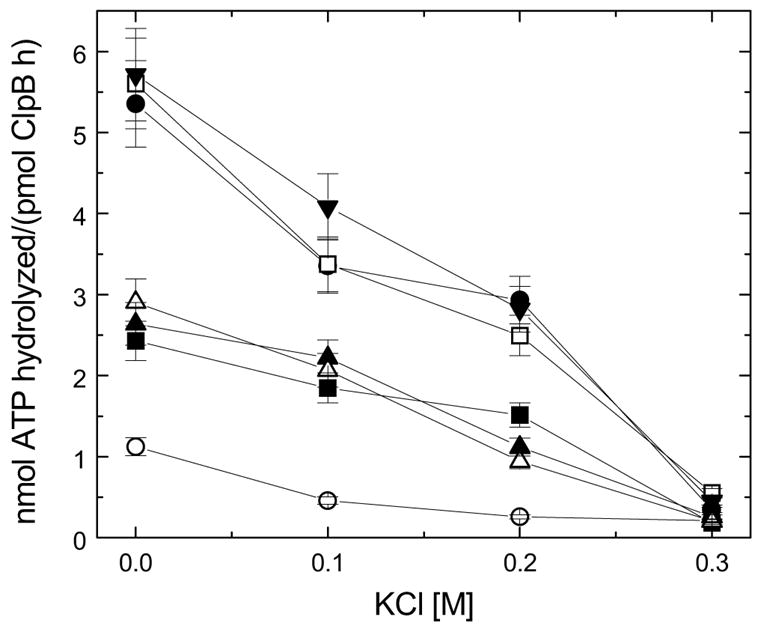
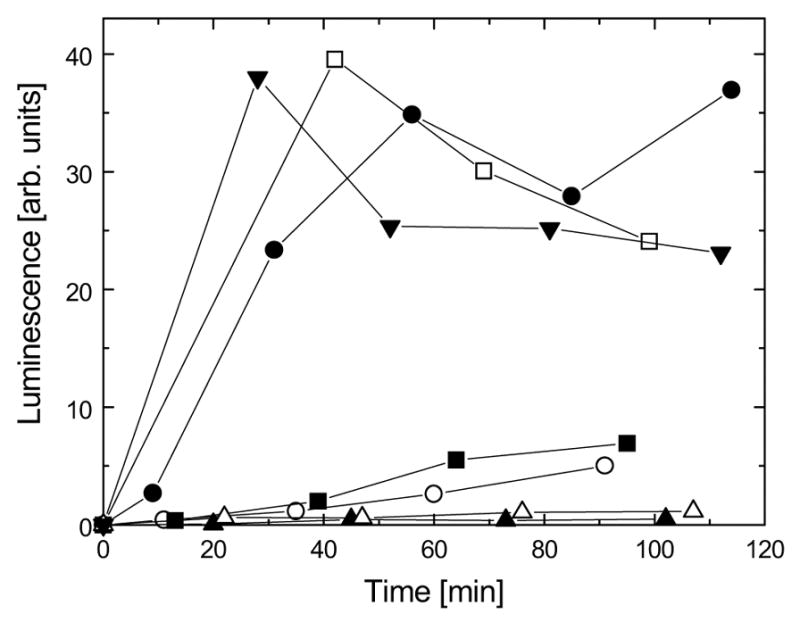
ClpB is a member of a multichaperone system in Escherichia coli (with DnaK, DnaJ, and GrpE) that reactivates strongly aggregated proteins. The sequence of ClpB contains two ATP-binding domains, each containing Walker consensus motifs. The N- and C-terminal sequence regions of ClpB do not contain known functional motifs. In this study, we performed site-directed mutagenesis of selected charged residues within the Walker A motifs (Lys212 and Lys611) and the C-terminal region of ClpB (Asp797, Arg815, Arg819, and Glu826). We found that the mutations K212T, K611T, D797A, R815A, R819A, and E826A did not significantly affect the secondary structure of ClpB. The mutation of the N-terminal ATP-binding site (K212T), but not of the C-terminal ATP-binding site (K611T), and two mutations within the C-terminal domain (R815A and R819A) inhibited the self-association of ClpB in the absence of nucleotides. The defects in self-association of these mutants were also observed in the presence of ATP and ADP. The four mutants K212T, K611T, R815A, and R819A showed an inhibition of chaperone activity, which correlated with their low ATPase activity in the presence of casein. Our results indicate that positively charged amino acids that are located along the intersubunit interface (this includes Lys212 in the Walker A motif of the N-terminal ATP-binding domain as well as Arg815 and Arg819 in the C-terminal domain) participate in intersubunit salt bridges and stabilize the ClpB oligomer. Interestingly, we have identified a conserved residue within the C-terminal domain (Arg819) which does not participate directly in nucleotide binding but is essential for the chaperone activity of ClpB.
Figures




References
-
- Schirmer EC, Glover JR, Singer MA, Lindquist S. Trends Biochem Sci. 1996;21:289–296. - PubMed
-
- Glover JR, Tkach JM. Biochem Cell Biol. 2001;79:557–568. - PubMed
-
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. Genome Res. 1999;9:27–43. - PubMed
-
- Weber-Ban EU, Reid BG, Miranker AD, Horwich AL. Nature. 1999;401:90–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
