

Calmodulin is an auxiliary subunit of KCNQ2/3 potassium channels
- PMID: 12223552
- PMCID: PMC6758071
- DOI: 10.1523/JNEUROSCI.22-18-07991.2002
Calmodulin is an auxiliary subunit of KCNQ2/3 potassium channels
Abstract
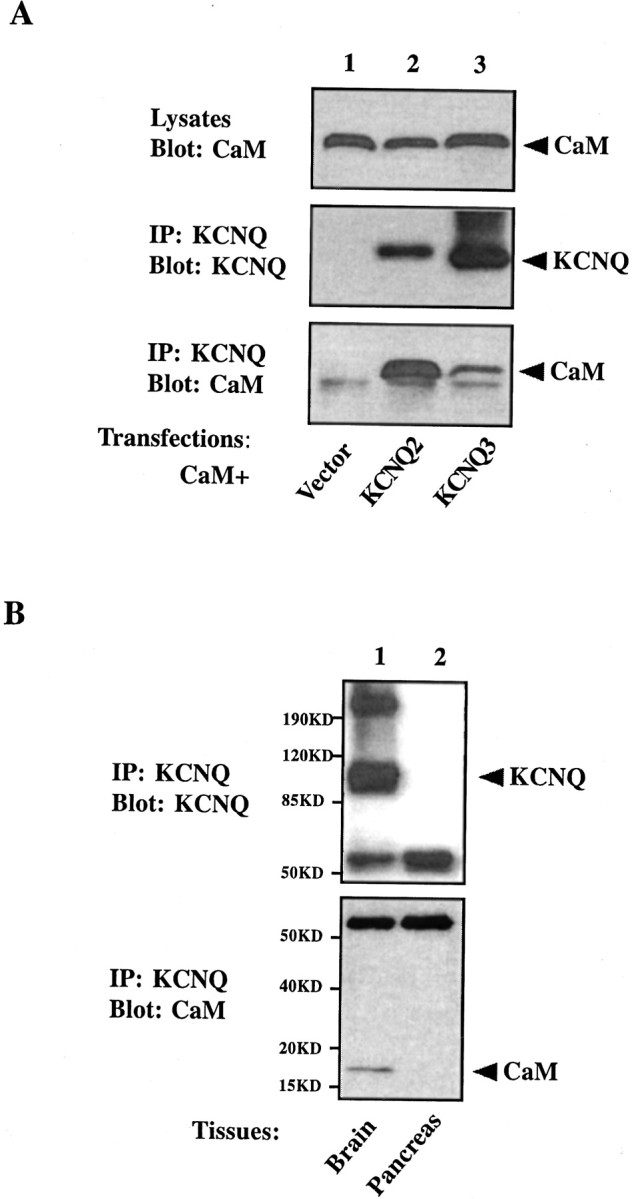
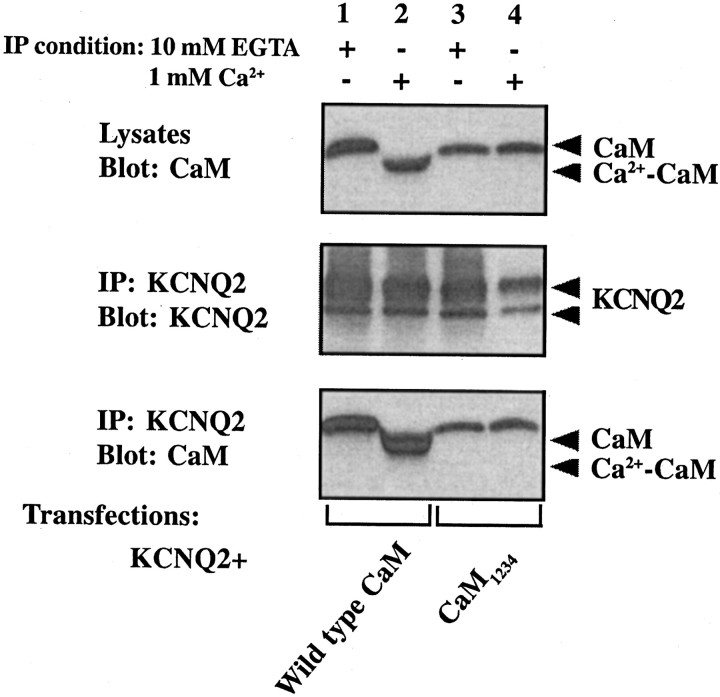
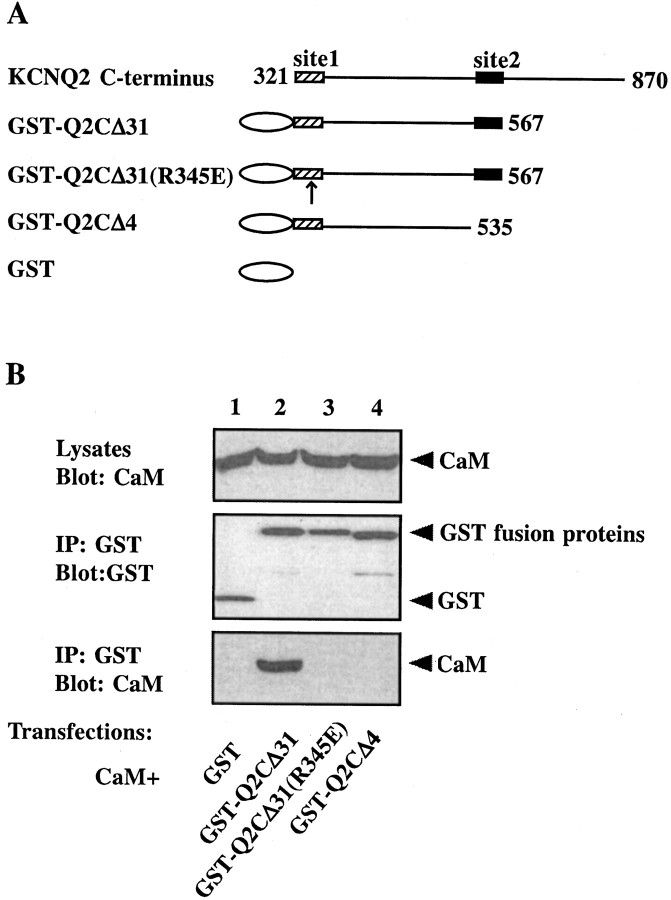
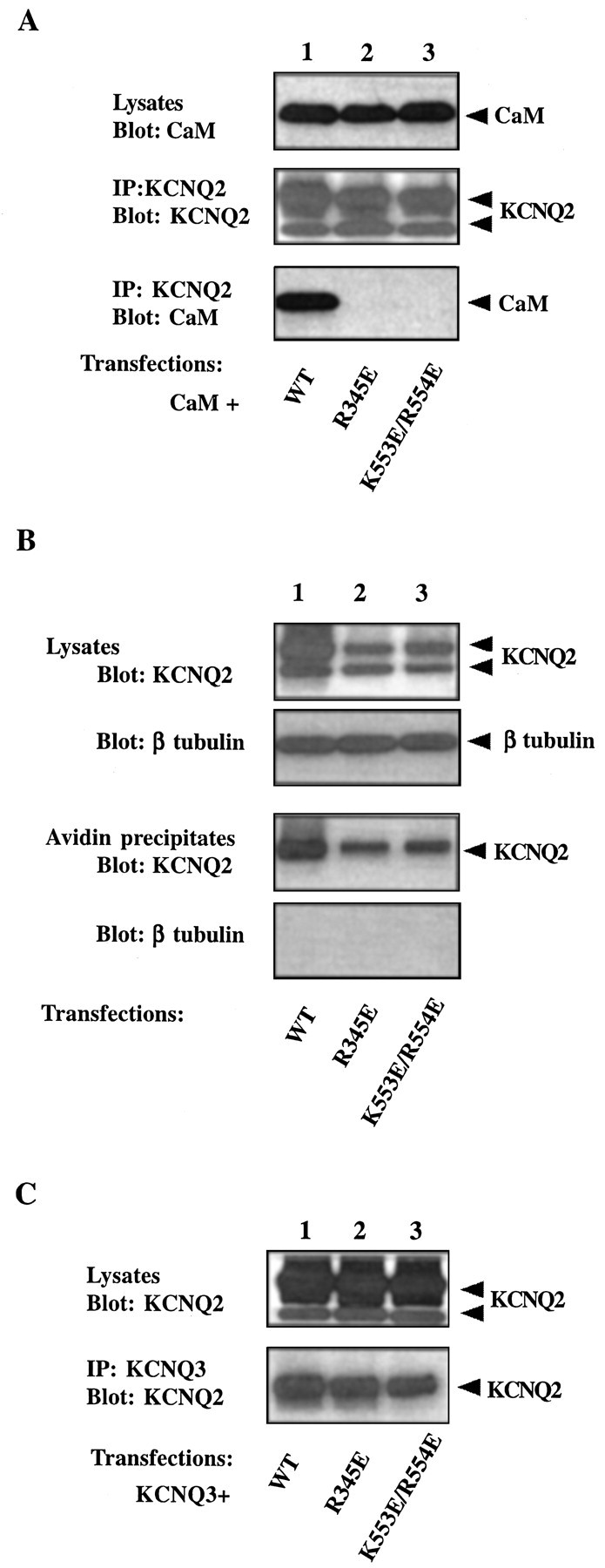
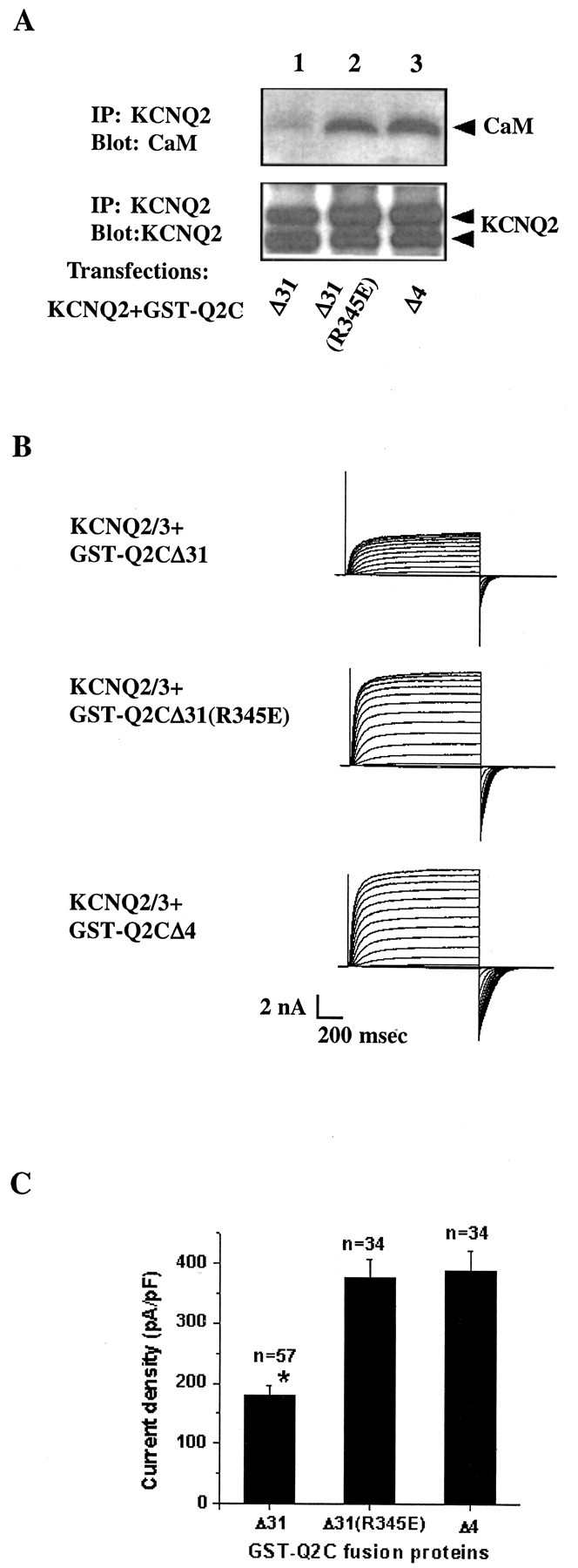
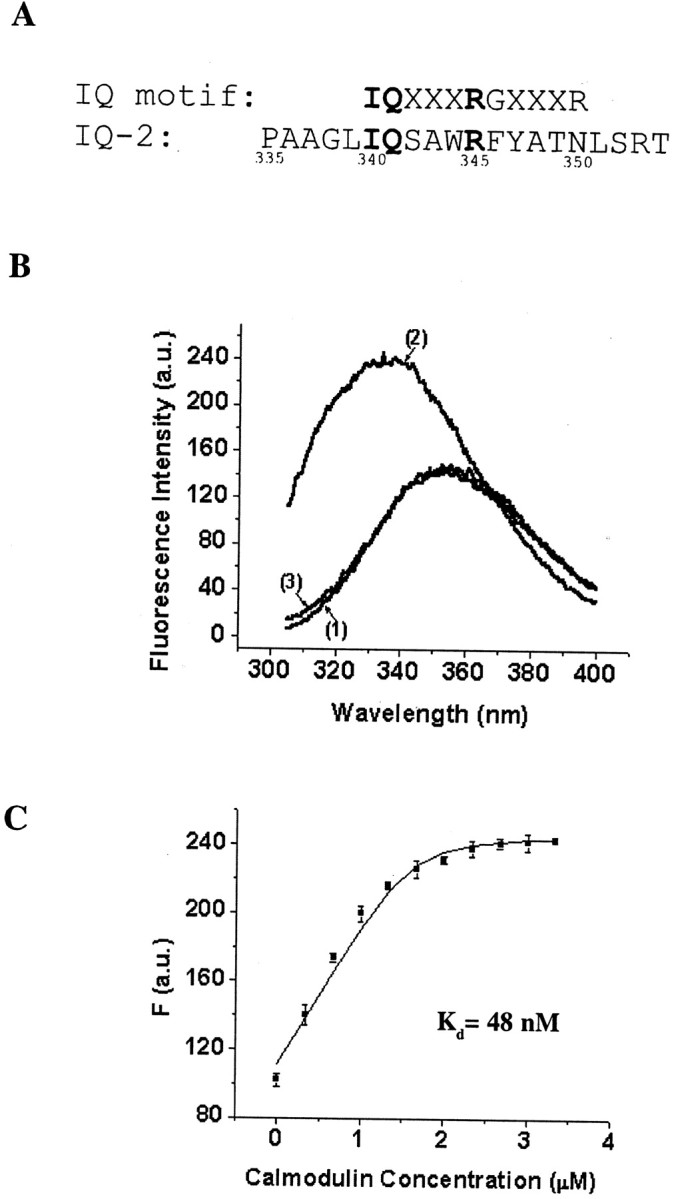
Calmodulin (CaM) was identified as a KCNQ2 and KCNQ3 potassium channel-binding protein, using a yeast two-hybrid screen. CaM is tethered constitutively to the channel, in the absence or presence of Ca2+, in transfected cells and also coimmunoprecipitates with KCNQ2/3 from mouse brain. The structural elements critical for CaM binding to KCNQ2 lie in two conserved motifs in the proximal half of the channel C-terminal domain. Truncations and point mutations in these two motifs disrupt the interaction. The first CaM-binding motif has a sequence that conforms partially to the consensus IQ motif, but both wild-type CaM and a Ca2+-insensitive CaM mutant bind to KCNQ2. The voltage-dependent activation of the KCNQ2/3 channel also shows no Ca2+ sensitivity, nor is it affected by overexpression of the Ca2+-insensitive CaM mutant. On the other hand, KCNQ2 mutants deficient in CaM binding are unable to generate detectable currents when coexpressed with KCNQ3 in CHO cells, although they are expressed and targeted to the cell membrane and retain the ability to assemble with KCNQ3. A fusion protein containing both of the KCNQ2 CaM-binding motifs competes with the full-length KCNQ2 channel for CaM binding and decreases KCNQ2/3 current density in CHO cells. The correlation of CaM binding with channel function suggests that CaM is an auxiliary subunit of the KCNQ2/3 channel.
Figures

References
-
- Biervert C, Schroeder BC, Kubisch C, Berkovic SF, Propping P, Jentsch TJ, Steinlein OK. A potassium channel mutation in neonatal human epilepsy. Science. 1998;279:403–406. - PubMed
-
- Brown DA, Adams PR. Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. Nature. 1980;283:673–676. - PubMed
-
- Charlier C, Singh NA, Ryan SG, Lewis TB, Reus BE, Leach RJ, Leppert M. A pore mutation in a novel KQT-like potassium channel gene in an idiopathic epilepsy family. Nat Genet. 1998;18:53–55. - PubMed
-
- El Far O, Bofill-Cardona E, Airas JM, O'Connor V, Boehm S, Freissmuth M, Nanoff C, Betz H. Mapping of calmodulin and G-βγ binding domains within the C-terminal region of the metabotropic glutamate receptor 7A. J Biol Chem. 2001;276:30662–30669. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous