

Computational discovery of sense-antisense transcription in the human and mouse genomes
- PMID: 12225583
- PMCID: PMC126869
- DOI: 10.1186/gb-2002-3-9-research0044
Computational discovery of sense-antisense transcription in the human and mouse genomes
Abstract
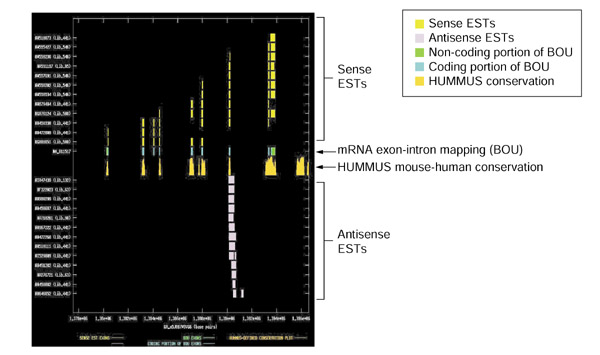
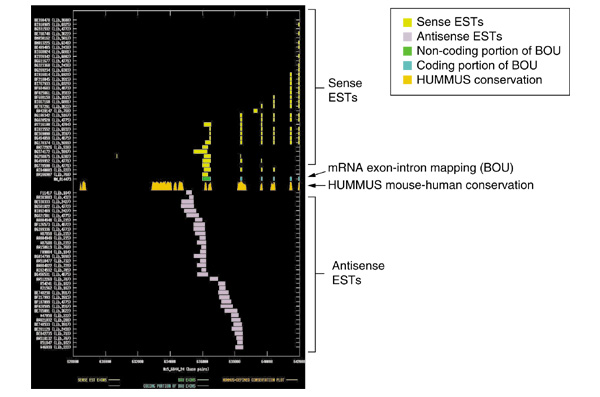
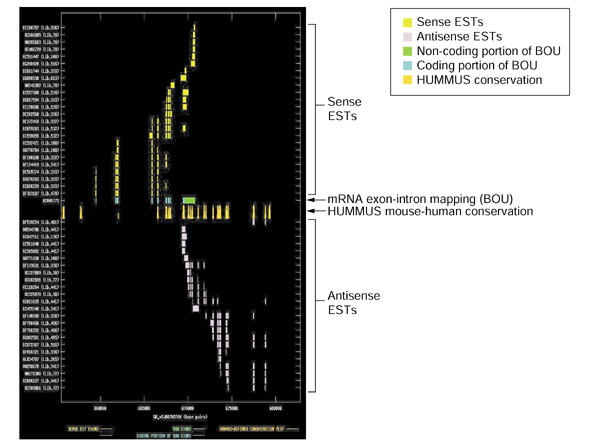
Background: Overlapping but oppositely oriented transcripts have the potential to form sense-antisense perfect double-stranded (ds) RNA duplexes. Over recent years, the number and variety of examples of mammalian gene-regulatory phenomena in which endogenous dsRNA duplexes have been proposed or demonstrated to participate has greatly increased. These include genomic imprinting, RNA interference, translational regulation, alternative splicing, X-inactivation and RNA editing. We computationally mined public mouse and human expressed sequence tag (EST) databases to search for additional examples of bidirectionally transcribed genomic regions.
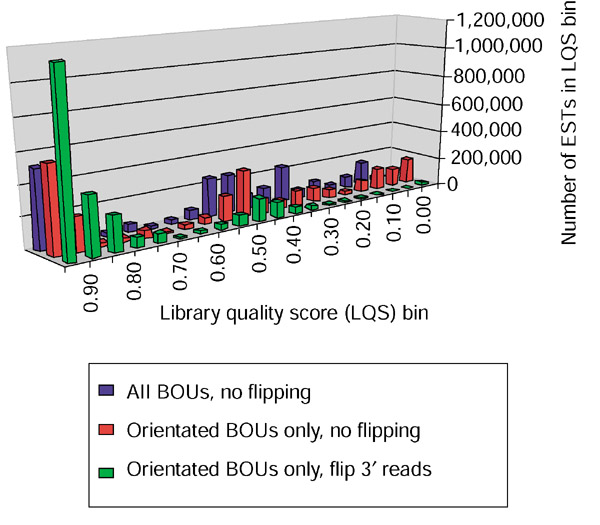
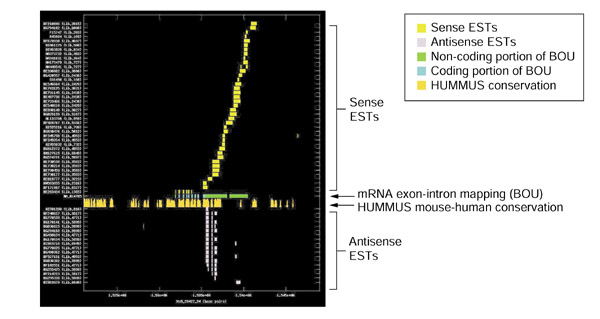
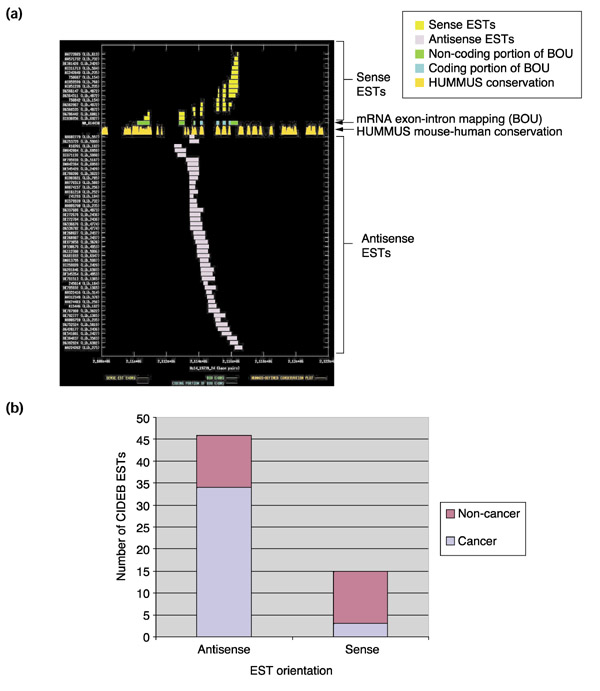
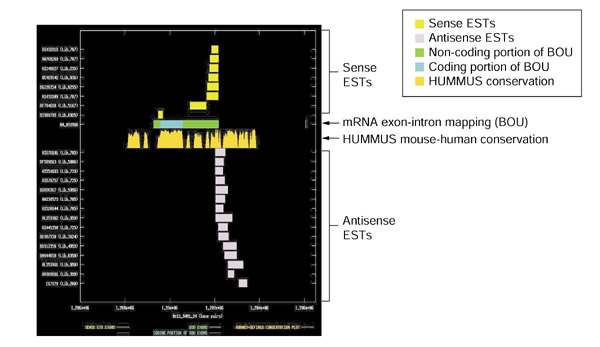
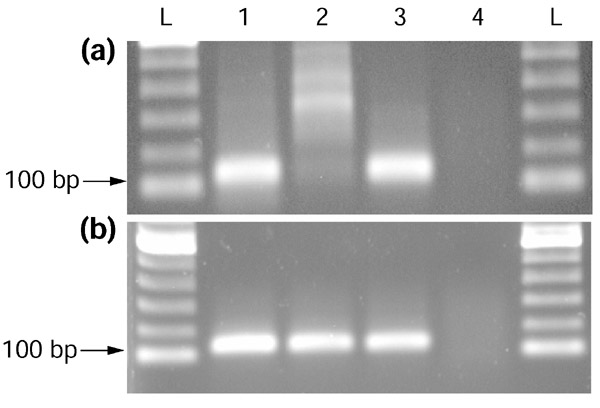
Results: Our bioinformatics approach identified over 217 candidate overlapping transcriptional units, almost all of which are novel. From experimental validation of a subset of our predictions by orientation-specific RT-PCR, we estimate that our methodology has a specificity of 84% or greater. In many cases, regions of sense-antisense overlap within the 5'- or 3'-untranslated regions of a given transcript correlate with genomic patterns of mouse-human conservation.
Conclusions: Our results, in conjunction with the literature, bring the total number of predicted and validated examples of overlapping but oppositely oriented transcripts to over 300. Several of these cases support the hypothesis that a subset of the instances of substantial mouse-human conservation in the 5' and 3' UTRs of transcripts might be explained in part by functionality of an overlapping transcriptional unit.
Figures
References
-
- Vanhee-Brossollet C, Vaquero C. Do natural antisense transcripts make sense in eukaryotes? Gene. 1998;211:1–9. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Sleutels F, Zwart R, Barlow DP. The non-coding Air RNA is required for silencing autosomal imprinted genes. Nature. 2002;415:810–813. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
