

Inositol phospholipid metabolism in Arabidopsis. Characterized and putative isoforms of inositol phospholipid kinase and phosphoinositide-specific phospholipase C
- PMID: 12226484
- PMCID: PMC166537
- DOI: 10.1104/pp.004770
Inositol phospholipid metabolism in Arabidopsis. Characterized and putative isoforms of inositol phospholipid kinase and phosphoinositide-specific phospholipase C
Abstract
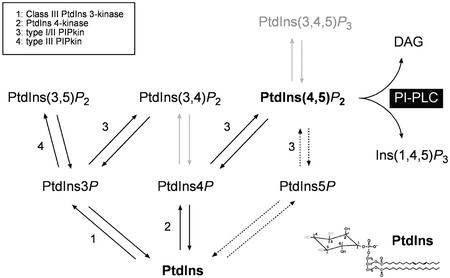
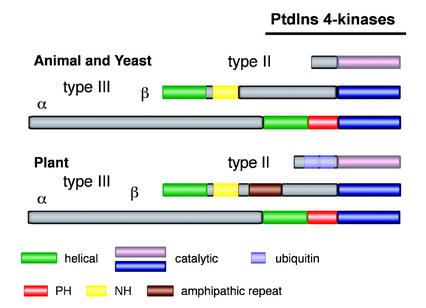
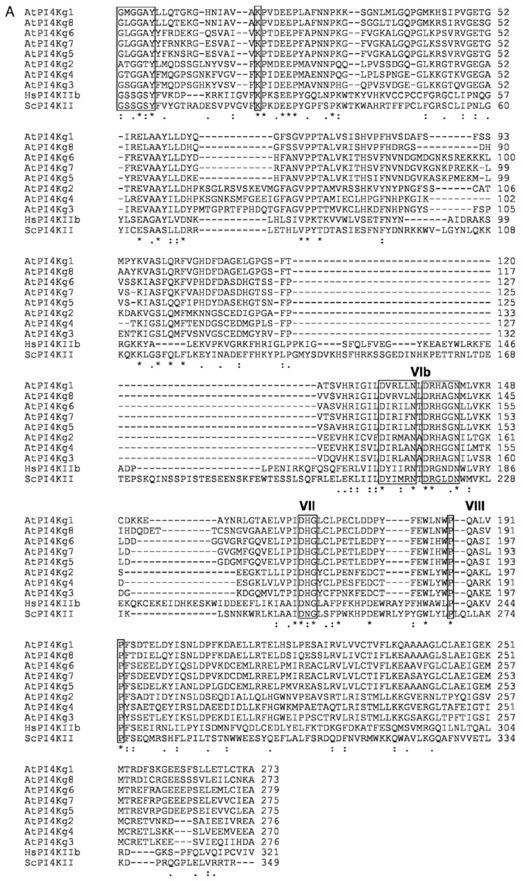
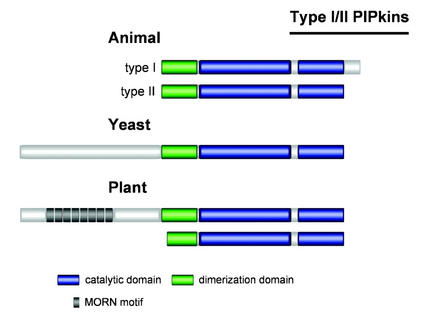
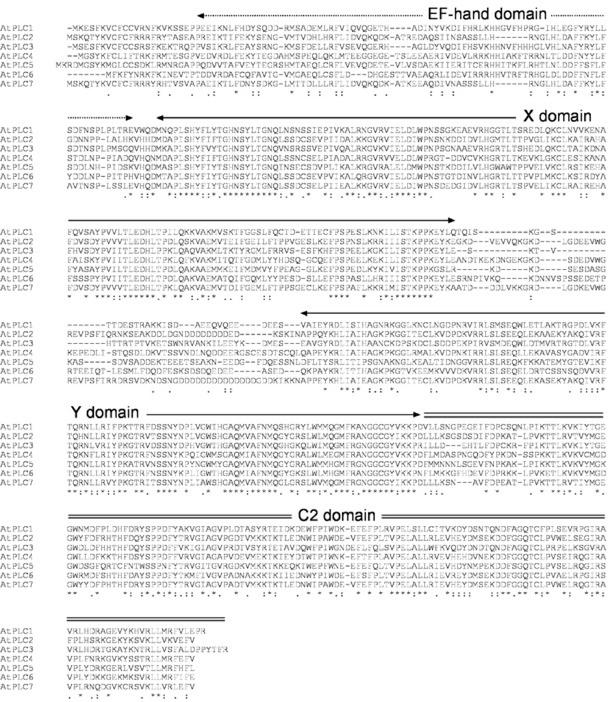
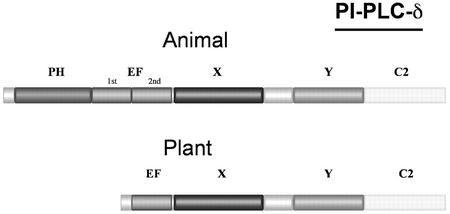
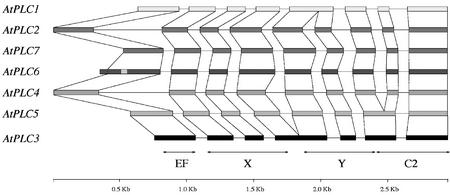
Phosphoinositides (PIs) constitute a minor fraction of total cellular lipids in all eukaryotic cells. They fulfill many important functions through interaction with a wide range of cellular proteins. Members of distinct inositol lipid kinase families catalyze the synthesis of these phospholipids from phosphatidylinositol. The hydrolysis of PIs involves phosphatases and isoforms of PI-specific phospholipase C. Although our knowledge of the roles played by plant PIs is clearly limited at present, there is no doubt that they are involved in many physiological processes during plant growth and development. In this review, we concentrate on inositol lipid-metabolizing enzymes from the model plant Arabidopsis for which biochemical characterization data are available, namely the inositol lipid kinases and PI-specific phospholipase Cs. The biochemical properties and structure of characterized and genome-predicted isoforms are presented and compared with those of the animal enzymes to show that the plant enzymes have some features clearly unique to this kingdom.
Figures



References
-
- Alexandre J, Lassalles JF, Kado RD. Opening of Ca2+channels in isolated red beet vacuole membrane by inositol 1,4,5-trisphosphate. Nature. 1990;343:567–570.
-
- Ananthanarayanan B, Das S, Rhee SG, Murray D, Cho W. Membrane targeting of C2 domains of phospholipase C-δ isoforms. J Biol Chem. 2001;277:3568–3575. - PubMed
-
- Bae YS, Cantley LG, Chen CS, Kim SR, Kwon KS, Rhee SG. Activation of phospholipase C-γ by phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem. 1998;273:4465–4469. - PubMed
-
- Balla T. Phosphatidylinositol 4-kinases. Biochim Biophys Acta. 1998;1436:69–85. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
