

Molecular identification of cytosolic, patatin-related phospholipases A from Arabidopsis with potential functions in plant signal transduction
- PMID: 12226489
- PMCID: PMC166542
- DOI: 10.1104/pp.006288
Molecular identification of cytosolic, patatin-related phospholipases A from Arabidopsis with potential functions in plant signal transduction
Abstract
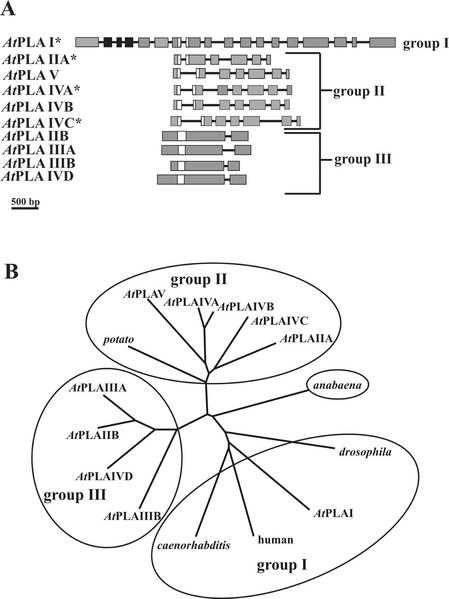
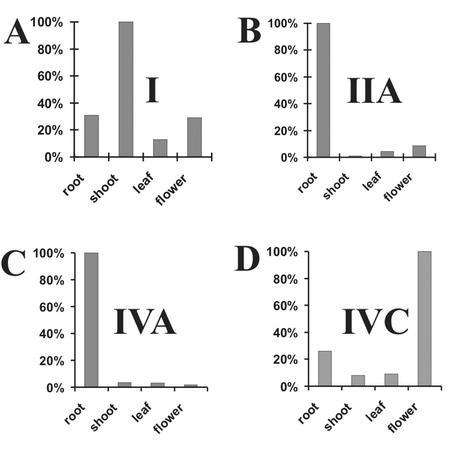
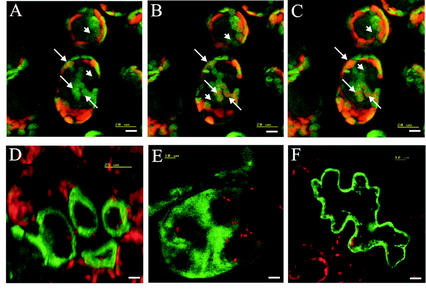
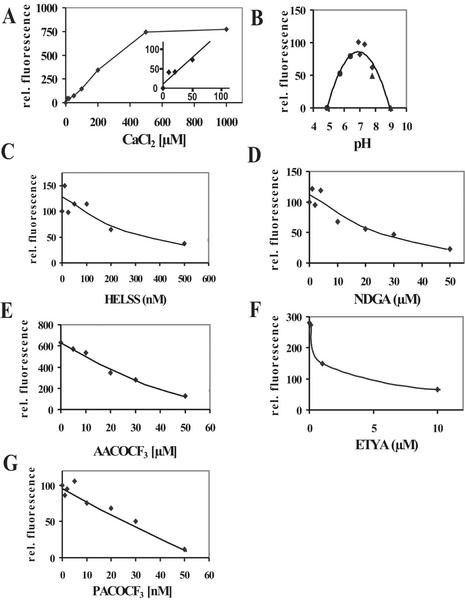
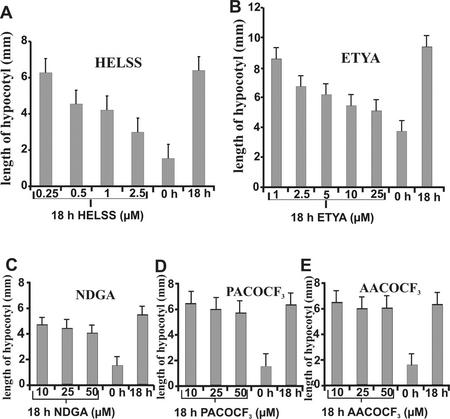
Rapid activation of phospholipase A (PLA) by auxin or plant-pathogen interaction suggests a function in signal transduction for this enzyme, but the molecular identification of a cytosolic PLA carrying out this function remains open. We isolated four cDNA sequences from Arabidopsis (ecotype Columbia), AtPLA I, AtPLA IIA, AtPLA IVA, and AtPLA IVC, which are members of the patatin-related PLA gene family in plants and which are homologous to the animal Ca(2+)-independent PLA(2) gene family. Expression was measured by reverse transcriptase-polymerase chain reaction, and AtPLA I transcripts were found preferentially in shoots, AtPLA IIA and AtPLA IVA in roots, and AtPLA IVC in flowers. Transient expression of the four PLA-green fluorescent protein fusion proteins in tobacco (Nicotiana tabacum) leaves showed they were located in the cytosol and not in the vacuoles. Surprisingly, AtPLA::green fluorescent protein was also localized to chloroplasts. The enzymatic activity of the purified recombinant AtPLA IVA toward phosphatidylcholine was dependent on Ca(2+), saturated at 0.5 mM, and had a pH optimum of about 7.0. It had both PLA(1) and PLA(2) specificity. The enzyme showed in vitro highest sensitivity toward the PLA(2) inhibitors palmitoyltrifluoromethyl ketone (PACOCF(3), K(i) approximately 30 nM), arachidonyltrifluoromethyl ketone (AACOCF(3), K(i) approximately 25 microM), and tetrahydro-3-(1-naphtalenyl)-2H-pyran-2-one (K(i) approximately 200 nM) and was also sensitive to other previously used inhibitors 5,8,11,14-eicosatetraynoic acid (K(i) approximately 3 microM) and nordihydroguajaretic acid (K(i) approximately 15 microM). The influence of these PLA(2) inhibitors on elongation in etiolated Arabidopsis seedlings was tested, and tetrahydro-3-(1-naphtalenyl)-2H-pyran-2-one and 5,8,11,14-eicosatetraynoic acid inhibited hypocotyl elongation maximally at concentrations close to their K(i) in vitro.
Figures


References
-
- Ackermann JE, Conde-Frieboes K, Dennis EA. Inhibition of macrophage Ca2+-independent phospholipase A2 by bromoenol lactone and trifluoromethyl ketones. J Biol Chem. 1995;270:445–450. - PubMed
-
- Balsinde J, Balboa MA, Insel PA, Dennis EA. Regulation and inhibition of phospholipase A2. Annu Rev Pharmacol Toxicol. 1999;39:175–189. - PubMed
-
- Balsinde J, Dennis EA. Function and inhibition of intracellular calcium-independent phospholipase A2. J Biol Chem. 1997;272:16069–16072. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
