

The divergently transcribed Streptococcus parasanguis virulence-associated fimA operon encoding an Mn(2+)-responsive metal transporter and pepO encoding a zinc metallopeptidase are not coordinately regulated
- PMID: 12228300
- PMCID: PMC128308
- DOI: 10.1128/IAI.70.10.5706-5714.2002
The divergently transcribed Streptococcus parasanguis virulence-associated fimA operon encoding an Mn(2+)-responsive metal transporter and pepO encoding a zinc metallopeptidase are not coordinately regulated
Abstract
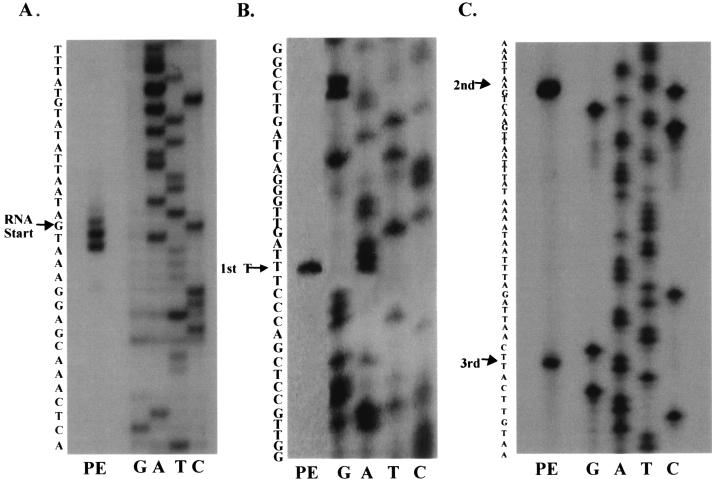
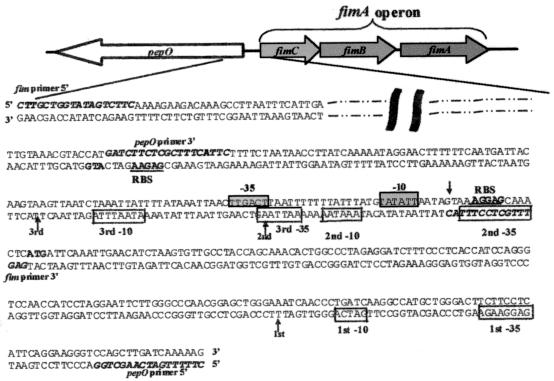
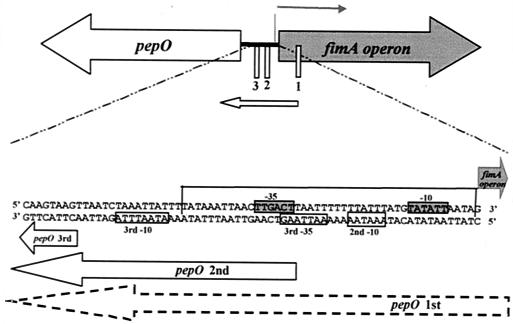
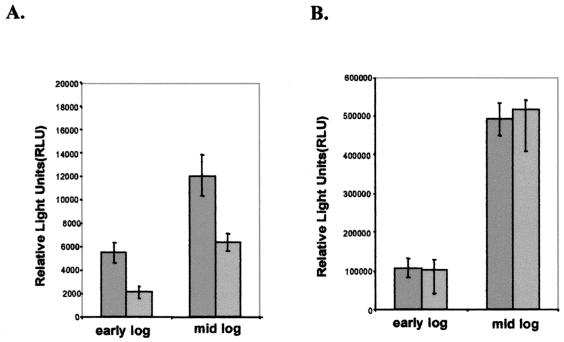
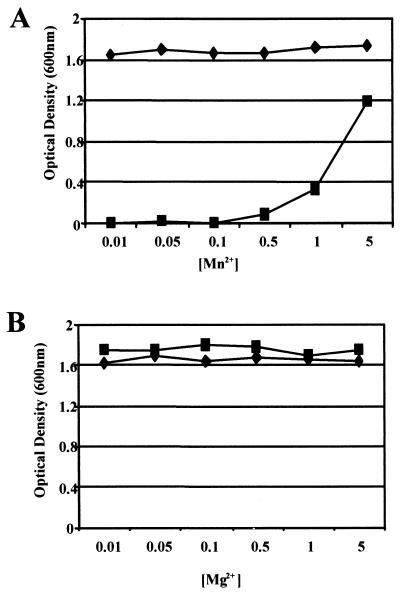
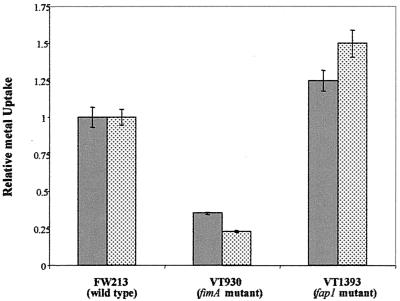
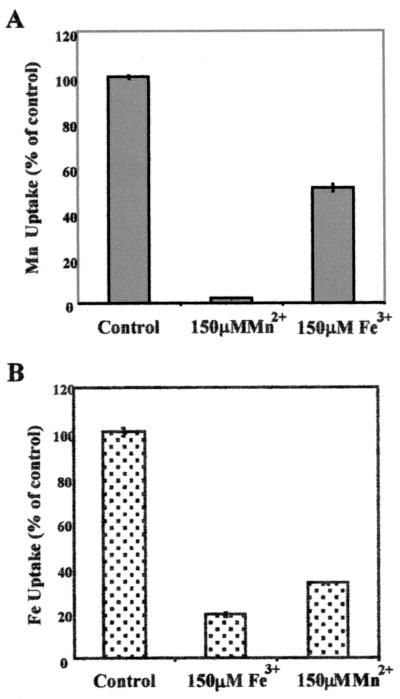
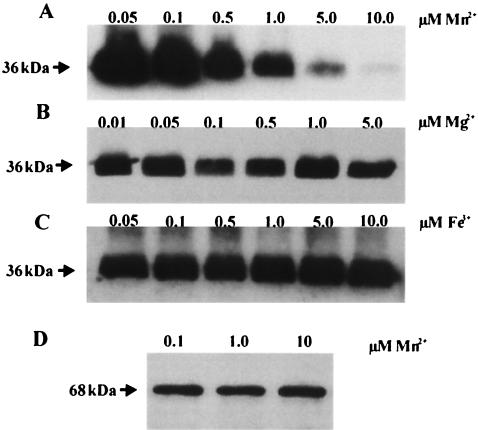
The study of how bacteria respond to and obtain divalent metal ions provides insight into the regulation of virulence factors in the host environment. Regulation of metal permease operons in gram-positive bacteria may involve the binding of metal-responsive repressors to palindromic domains in their control regions. The Streptococcus parasanguis fimA operon, which encodes an ATP-binding cassette (ABC) transporter system with sequence homology to the LraI family of metal transporters, possesses a palindromic regulatory region with high homology to that of the Streptococcus gordonii ScaR binding domain. Mapping of the promoter and regulatory regions of fimA and the divergently transcribed pepO gene, which encodes a zinc metalloendopeptidase, indicated that their promoter and regulatory elements overlap. fimA had one transcriptional start site, whereas pepO had three. Analysis of truncated versions of the pepO promoter suggested that all three transcriptional start sites are functional. Analysis of promoter activity under various environmental conditions indicated that the fimA operon promoter and the pepO promoter are not coordinately regulated. The fimA operon is responsive to changes in Mn(2+) concentration, but the pepO promoter is not. A S. parasanguis fimA mutant showed a growth deficiency under conditions of limiting Mn(2+). This deficiency was not alleviated by compensation with either Mg(2+) or Fe(3+). Wild-type S. parasanguis could take up Mn(2+) and Fe(3+), while the fimA mutant showed a marked reduction in this ability. These data suggested that FimA is a component of a metal transporter system capable of transporting both Mn(2+) and Fe(3+). FimA expression itself was shown to be responsive to Mn(2+) concentration, but not to availability of Fe(3+) or Mg(2+).
Figures
Similar articles
-
Streptococcus parasanguis pepO encodes an endopeptidase with structure and activity similar to those of enzymes that modulate peptide receptor signaling in eukaryotic cells.Infect Immun. 1999 Oct;67(10):5206-14. doi: 10.1128/IAI.67.10.5206-5214.1999. Infect Immun. 1999. PMID: 10496897 Free PMC article.
-
The fimA locus of Streptococcus parasanguis encodes an ATP-binding membrane transport system.Mol Microbiol. 1995 Mar;15(5):849-63. doi: 10.1111/j.1365-2958.1995.tb02355.x. Mol Microbiol. 1995. PMID: 7596287
-
Expression of the virulence-related Sca (Mn2+) permease in Streptococcus gordonii is regulated by a diphtheria toxin metallorepressor-like protein ScaR.Mol Microbiol. 2000 Oct;38(1):140-53. doi: 10.1046/j.1365-2958.2000.02122.x. Mol Microbiol. 2000. PMID: 11029696
-
Competence and virulence of Streptococcus pneumoniae: Adc and PsaA mutants exhibit a requirement for Zn and Mn resulting from inactivation of putative ABC metal permeases.Mol Microbiol. 1997 Aug;25(4):727-39. doi: 10.1046/j.1365-2958.1997.5111879.x. Mol Microbiol. 1997. PMID: 9379902
-
Manganese transport and the role of manganese in virulence.Annu Rev Microbiol. 2006;60:187-209. doi: 10.1146/annurev.micro.60.080805.142149. Annu Rev Microbiol. 2006. PMID: 16704341 Review.
Cited by
-
PepO, a CovRS-controlled endopeptidase, disrupts Streptococcus pyogenes quorum sensing.Mol Microbiol. 2016 Jan;99(1):71-87. doi: 10.1111/mmi.13216. Epub 2015 Oct 14. Mol Microbiol. 2016. PMID: 26418177 Free PMC article.
-
Biology of Oral Streptococci.Microbiol Spectr. 2018 Oct;6(5):10.1128/microbiolspec.gpp3-0042-2018. doi: 10.1128/microbiolspec.GPP3-0042-2018. Microbiol Spectr. 2018. PMID: 30338752 Free PMC article. Review.
-
Manganese uptake and streptococcal virulence.Biometals. 2015 Jun;28(3):491-508. doi: 10.1007/s10534-015-9826-z. Epub 2015 Feb 5. Biometals. 2015. PMID: 25652937 Free PMC article. Review.
-
Manganese acquisition is essential for virulence of Enterococcus faecalis.PLoS Pathog. 2018 Sep 20;14(9):e1007102. doi: 10.1371/journal.ppat.1007102. eCollection 2018 Sep. PLoS Pathog. 2018. PMID: 30235334 Free PMC article.
-
The sloABCR operon of Streptococcus mutans encodes an Mn and Fe transport system required for endocarditis virulence and its Mn-dependent repressor.J Bacteriol. 2003 Oct;185(20):5967-75. doi: 10.1128/JB.185.20.5967-5975.2003. J Bacteriol. 2003. PMID: 14526007 Free PMC article.
References
-
- Andersen, R. N., R. D. Lunsford, and P. E. Kolenbrander. 1997. Determination of the transcript size and start site of the putative sca operon of Streptococcus gordonii ATCC 51656 (formerly strain PK488). Adv. Exp. Med. Biol. 418:657-660. - PubMed
-
- Ausubel, F. M. 1988. Current protocols in molecular biology. Greene Publishers and Associates and Wiley-Interscience, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
