

doi: 10.1101/gad.1015202.
Autophosphorylation of the DNA-dependent protein kinase catalytic subunit is required for rejoining of DNA double-strand breaks
Affiliations
- PMID: 12231622
- PMCID: PMC187438
- DOI: 10.1101/gad.1015202
Item in Clipboard
Autophosphorylation of the DNA-dependent protein kinase catalytic subunit is required for rejoining of DNA double-strand breaks
Genes Dev.
.
Abstract
Nonhomologous end-joining (NHEJ) is the predominant pathway that repairs DNA double-strand breaks (DSBs) in mammalian cells. The DNA-dependent protein kinase (DNA-PK), consisting of Ku and DNA-PK catalytic subunit (DNA-PKcs), is activated by DNA in vitro and is required for NHEJ. We report that DNA-PKcs is autophosphorylated at Thr2609 in vivo in a Ku-dependent manner in response to ionizing radiation. Phosphorylated DNA-PKcs colocalizes with both gamma-H2AX and 53BP1 after DNA damage. Mutation of Thr2609 to Ala leads to radiation sensitivity and impaired DSB rejoining. These findings establish that Ku-dependent phosphorylation of DNA-PKcs at Thr2609 is required for the repair of DSBs by NHEJ.
Figures
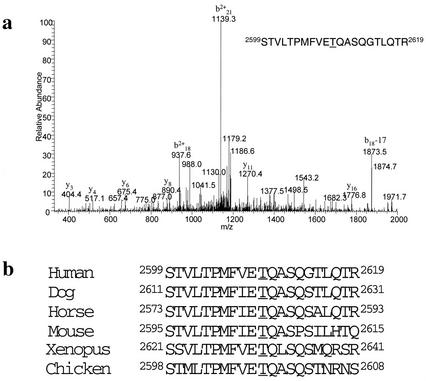
DNA-PKcs phosphorylation of Thr2609. (a) Mass spectrometry (MS) identification of the DNA-PKcs in vitro autophosphorylation site. MS spectra of the identified tryptic-phosphopeptide; the underlined T corresponds to Thr2609 and is the site of phosphorylation. (b) Alignment of human DNA-PKcs Thr2609 with DNA-PKcs sequences available in the National Center for Biotechnology Information database.
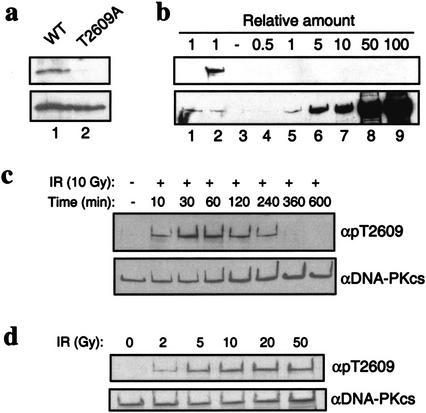
Characterization of Thr2609 phosphorylation. (a) Specificity of pT2609Ab. GST fragments spanning amino acids 2500–2700 that contain either the wild-type DNA-PKcs sequence or a T2609A point mutation were in vitro phosphorylated with purified DNA-PK and analyzed with the pT2609Ab (top) and anti-GST (bottom) to show equal loading. (b) pT2609Ab does not recognize unphosphorylated DNA-PKcs. Western blotting with pT2609Ab (top) and 25-4 DNA-PKcs monoclonal antibody (bottom) with mock or autophosphorylated DNA-PKcs (lanes 1 and 2, respectively) and with purified, unphosphorylated DNA-PKcs (lanes 3–9) at the indicated molar ratios relative to the amount of protein in lanes 1,2. (c) Kinetics of Thr2609 phosphorylation in vivo. HeLa cells were either mock-treated or irradiated with 10 Gy and allowed to recover for the indicated times. Nuclear extracts were Western blotted with pT2609Ab (top), and then blots were stripped and reprobed with the 25-4 DNA-PKcs monoclonal antibody (bottom). (d) Dose dependence of Thr2609 phosphorylation. HeLa cells were irradiated with the indicated dose of ionizing radiation and allowed to recover for 30 min. Nuclear extracts were first analyzed by Western blot with pT2609Ab (top) and then with 25-4 monoclonal to show equal loading (bottom).
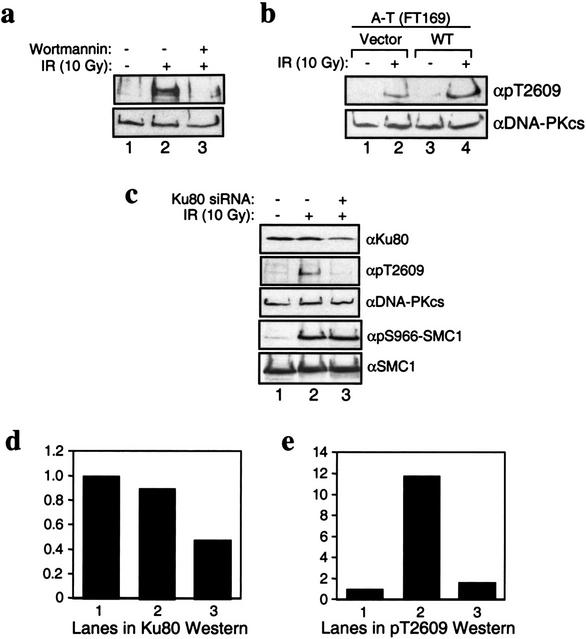
Characterization of Thr2609 phosphorylation in vivo. (a) Inhibition of Thr2609 phosphorylation by wortmannin. HeLa cells were preincubated with DMSO (d im ethyl s ulfo xide; lanes 1,2) or 20 μM wortmannin (lane 3) for 30 min before treatment with 10 Gy of ionizing radiation (IR; lanes 2,3). Nuclear extracts were prepared 30 min after IR and subsequently analyzed for Thr2609 phosphorylation (top) and with 25-4 monoclonal (bottom). (b) Phosphorylation of Thr2609 is ATM-independent. A-T cells (FT169) complemented with vector (lanes 1,2) or full-length wild-type ATM cDNA (lanes 3,4) were treated with 10 Gy of IR. Nuclear extracts were analyzed for Thr2609 phosphorylation (top) and DNA-PKcs levels (bottom). (c) Phosphorylation of Thr2609 is Ku-dependent. HeLa cells were mock-transfected (lanes 1,2) or transfected with a short interfering Ku80 RNA (lane 3) for 96 h before irradiation with 10 Gy of IR. Nuclear extracts were analyzed by Western blotting with the indicated antibodies. (d) Quantification of Ku80 Western signal from c. Values reported are relative to the signal observed in lane 1 and assigned an arbitrary value of one. (e) Quantification of pT2609Ab signal from c. Values reported are relative to the signal observed in lane 1.
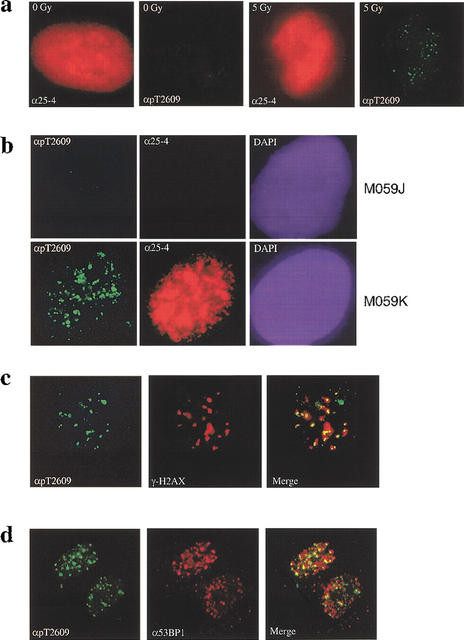
DNA-PKcs forms foci in response to ionizing radiation (IR). (a) DNA damage-induced pT2609 foci. Primary human fibroblasts were mock-treated or irradiated with 5 Gy of IR (as indicated) and allowed to recover for 30 min before fixation and staining with 25-4 monoclonal to DNA-PKcs (red) and pT2609Ab (green). (b) pT2609 foci are absent in DNA-PKcs-deficient cells. The DNA-PKcs-deficient (M059J) and the DNA-PKcs-proficient (M059K) cell lines were irradiated with 5 Gy and allowed to recover for 30 min before immunostaining with 25-4 monoclonal (red) and pT2609Ab (green) antibodies. (c) Colocalization of phosphorylated DNA-PKcs with γ-H2AX. Human primary fibroblasts were irradiated with 1 Gy, fixed at 30 min after IR, and stained with pT2609Ab (green) and monoclonal antibody against phosphoSer139 of histone H2AX (red; Upstate Biotech). The two images were merged, and colocalization of pT2609 and γ-H2AX foci are depicted as yellow. (d) Colocalization of phosphorylated DNA-PKcs with 53BP1. The experiment was conducted as in c but using 53BP1 monoclonal (red).
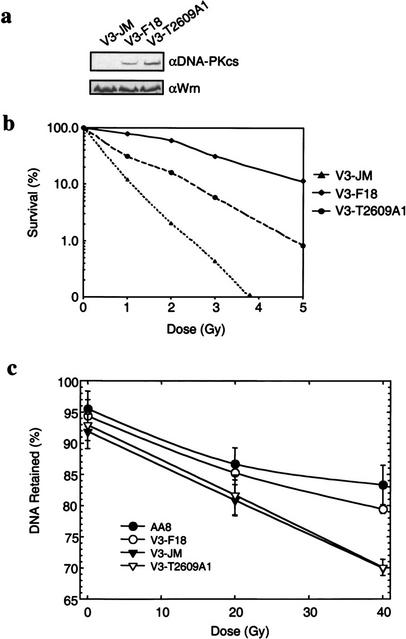
Phosphorylation of Thr2609 is required for double-strand break (DSB) repair. (a) Protein expression levels in V3-complemented cells. Nuclear extracts prepared from V3 transfected with vector alone (V3-JM), full-length wild-type DNA-PKcs (V3-F18), or DNA-PKcs containing the T2609A point mutant (V3-T2609A1) were analyzed for DNA-PKcs protein expression levels (top). Hamster Werner proteins were analyzed to show equal sample loading (bottom). (b) The colony formation assay was performed to compare radiation sensitivity of the three transfected V3 cell lines. V3-JM, V3-F18, and V3-T2609A1 were irradiated at the indicated doses and plated for analysis of survival and colony-forming ability. (c) FAR (f raction of a ctivity r eleased) assay to measure the presence of DNA DSBs. The V3-JM, V3-F18, and V3-T2609A1 cell lines and the parental CHO cell line (AA8) were irradiated at the indicated dose and analyzed for the presence of DSBs by the FAR assay. The greater the percentage of DNA retained, the fewer the number of DSBs remained.
References
-
- Banin S, Moyal L, Shieh S, Taya Y, Anderson CW, Chessa L, Smorodinsky NI, Prives C, Reiss Y, Shiloh Y, et al. Enhanced phosphorylation of p53 by ATM in response to DNA damage. Science. 1998;281:1674–1677. - PubMed
-
- Burma S, Chen BP, Murphy M, Kurimasa A, Chen DJ. ATM phosphorylates histone H2AX in response to DNA double-strand breaks. J Biol Chem. 2001;276:42462–42467. - PubMed
-
- Canman CE, Lim DS, Cimprich KA, Taya Y, Tamai K, Sakaguchi K, Appella E, Kastan MB, Siliciano JD. Activation of the ATM kinase by ionizing radiation and phosphorylation of p53. Science. 1998;281:1677–1679. - PubMed
-
- Chan DW, Lees-Miller SP. The DNA-dependent protein kinase is inactivated by autophosphorylation of the catalytic subunit. J Biol Chem. 1996;271:8936–8941. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous