

Visualization of parental HSV-1 genomes and replication compartments in association with ND10 in live infected cells
- PMID: 12234938
- PMCID: PMC126275
- DOI: 10.1093/emboj/cdf458
Visualization of parental HSV-1 genomes and replication compartments in association with ND10 in live infected cells
Abstract
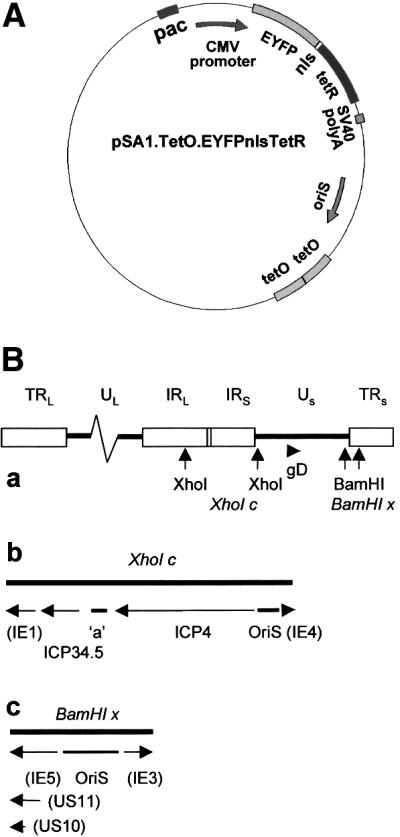
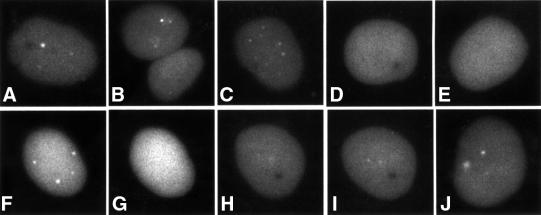
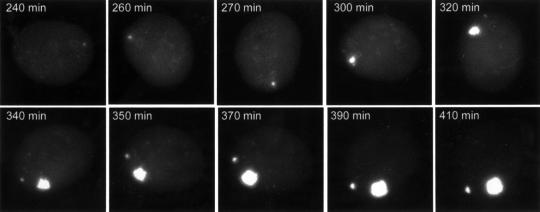
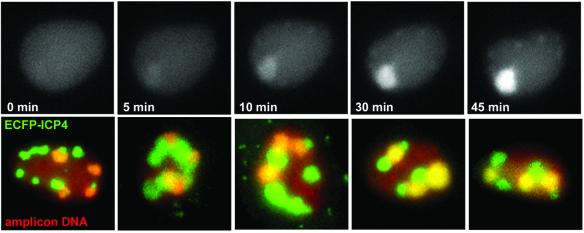
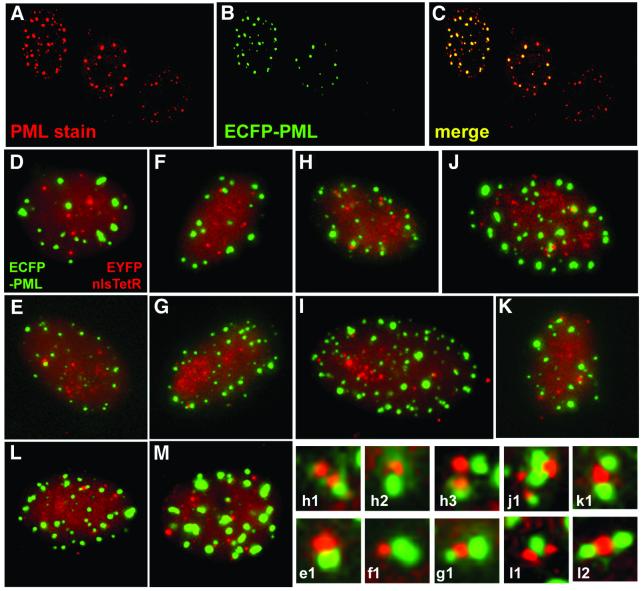
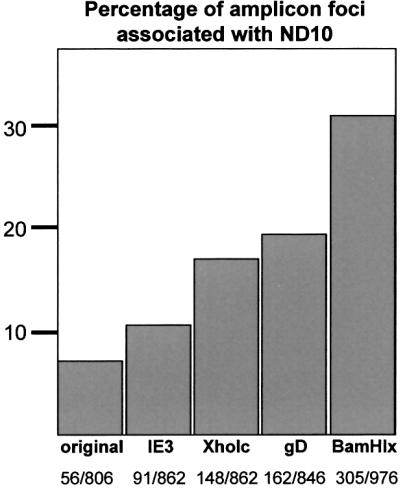
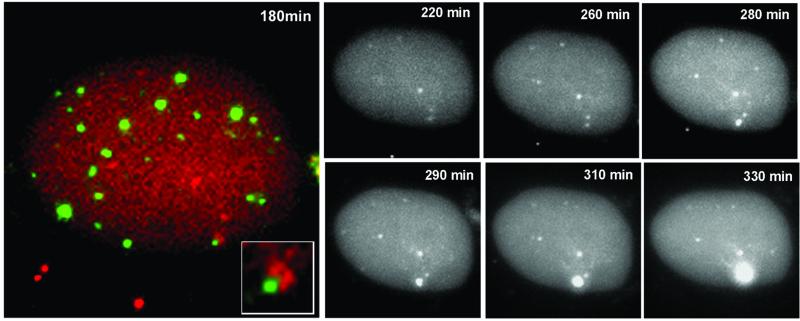
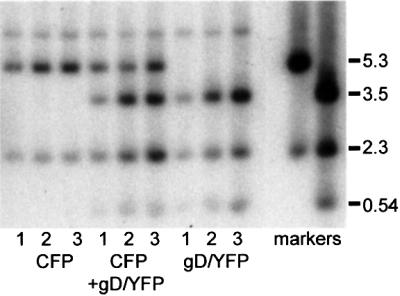
The relative location of active and repressed genes within the nucleus is becoming recognized as a significant factor in the control of gene expression. We have developed systems to visualize parental and replicated herpes simplex virus type 1 (HSV-1) amplicon genomes in association with PML nuclear bodies (ND10) in live cells. Plasmids containing viral replication and packaging signals, a gene expressing enhanced yellow fluorescent protein linked to the tetracycline repressor DNA binding domain and 14 copies of the tetracycline operator sequence were used to produce amplicon genomes packaged into normal viral particles. The frequency of the juxtaposition of viral genomes and ND10 was substantially increased by inclusion of an active HSV-1 Early gene transcription unit, indicating that the association is neither random nor passive. Furthermore, the ND10-associated genomes preferentially progressed to form viral replication compartments. Thus, active viral transcription contributes to the efficiency of viral genome association with ND10, and this in turn increases the probability that the genome will engage in active DNA replication. These studies in live cells provide a novel insight into virus-ND10 interactions and provide compelling visualization of their functional relevance.
Figures
References
-
- Belmont A.S. (2001) Visualizing chromosome dynamics with GFP. Trends Cell Biol., 11, 250–257. - PubMed
-
- Belmont A.S. and Straight,A.F. (1998) In vivo visualization of chromosomes using lac operator–repressor binding. Trends Cell Biol., 8, 121–124. - PubMed
-
- Cockell M. and Gasser,S.M. (1999) Nuclear compartments and gene regulation. Curr. Opin. Genet. Dev., 9, 199–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
