

Spontaneous access of proteins to buried nucleosomal DNA target sites occurs via a mechanism that is distinct from nucleosome translocation
- PMID: 12242292
- PMCID: PMC139820
- DOI: 10.1128/MCB.22.20.7147-7157.2002
Spontaneous access of proteins to buried nucleosomal DNA target sites occurs via a mechanism that is distinct from nucleosome translocation
Abstract
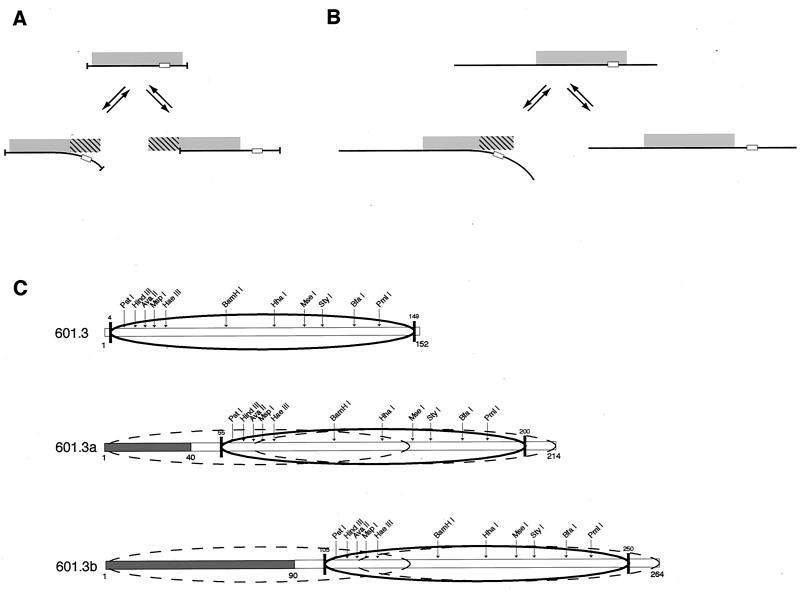
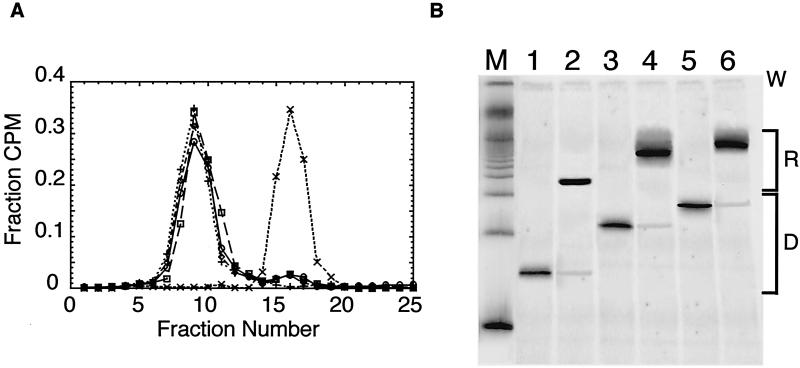
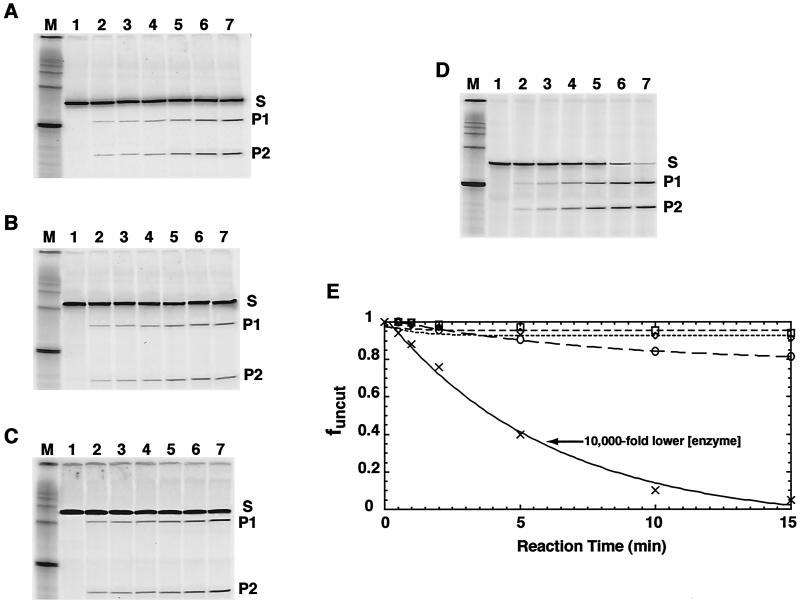
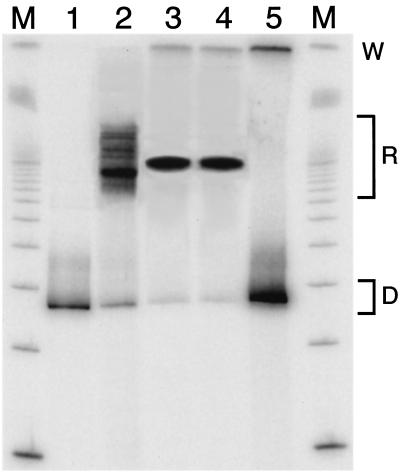
Intrinsic nucleosome dynamics termed "site exposure" provides spontaneous and cooperative access to buried regions of nucleosomal DNA in vitro. Two different mechanisms for site exposure have been proposed, one based on nucleosome translocation, the other on dynamic nucleosome conformational changes in which a stretch of the nucleosomal DNA is transiently released off the histone surface. Here we report on three experiments that distinguish between these mechanisms. One experiment investigates the effects on the accessibilities of restriction enzyme target sites inside nucleosomes when extra DNA (onto which the nucleosome may move at low energetic cost) is appended onto one end. The other two experiments test directly for nucleosome mobility under the conditions used to probe accessibility to restriction enzymes: one on a selected nonnatural nucleosome positioning sequence, the other on the well-studied 5S rRNA gene nucleosome positioning sequence. We find from all three assays that restriction enzymes gain access to sites throughout the entire length of the nucleosomal DNA without contribution from nucleosome translocation. We conclude that site exposure in nucleosomes in vitro occurs via a nucleosome conformational change that leads to transient release of a stretch of DNA from the histone surface, most likely involving progressive uncoiling from an end. Recapture at a distal site along DNA that has partially uncoiled would result in looped structures which are believed to contribute to RNA polymerase elongation and may contribute to spontaneous or ATP-driven nucleosome mobility. Transient open states may facilitate the initial entry of transcription factors and enzymes in vivo.
Figures


References
-
- Anderson, J. D., P. T. Lowary, and J. Widom. 2001. Effects of histone acetylation on the dynamic equilibrium accessibility of nucleosomal DNA target sites. J. Mol. Biol. 307:977-985. - PubMed
-
- Anderson, J. D., and J. Widom. 2000. Sequence- and position-dependence of the equilibrium accessibility of nucleosomal DNA target sites. J. Mol. Biol. 296:979-987. - PubMed
-
- Astumian, R. D., and M. Bier. 1994. Fluctuation driven ratchets: molecular motors. Phys. Rev. Lett. 72:1766-1769. - PubMed
-
- Bednar, J., V. M. Studitsky, S. A. Grigoryev, G. Felsenfeld, and C. L. Woodcock. 1999. The nature of the nucleosomal barrier to transcription: direct observation of paused intermediates by electron cryomicroscopy. Mol. Cell 4:377-386. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources