

Alpha-fetoprotein, the major fetal serum protein, is not essential for embryonic development but is required for female fertility
- PMID: 12297623
- PMCID: PMC130551
- DOI: 10.1073/pnas.202215399
Alpha-fetoprotein, the major fetal serum protein, is not essential for embryonic development but is required for female fertility
Abstract
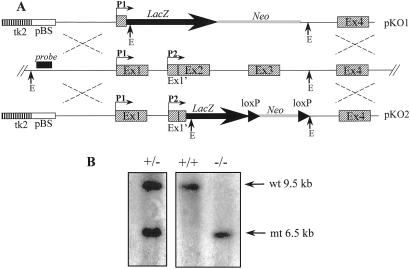
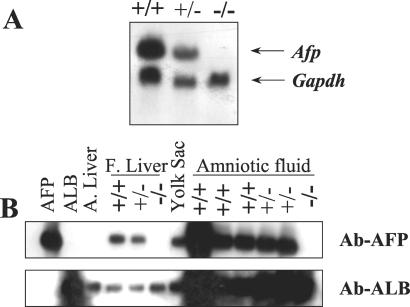
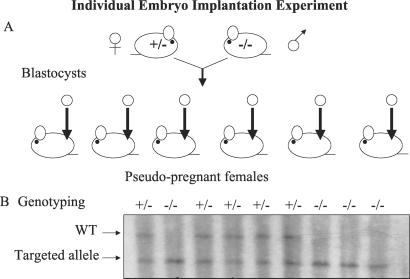
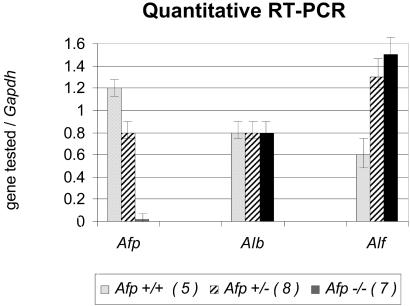
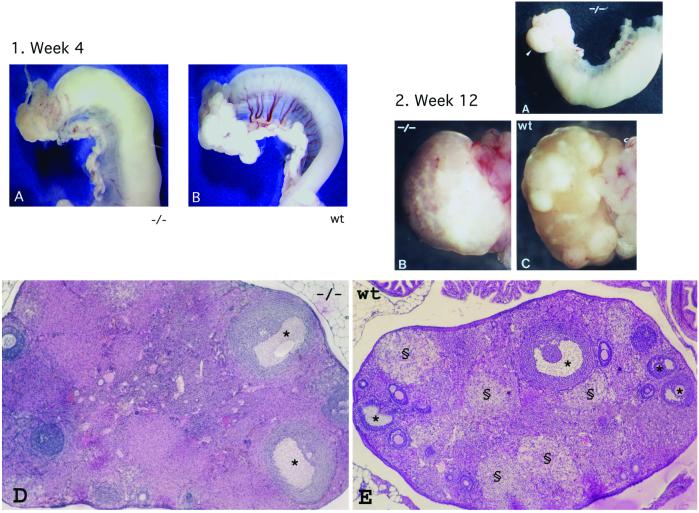
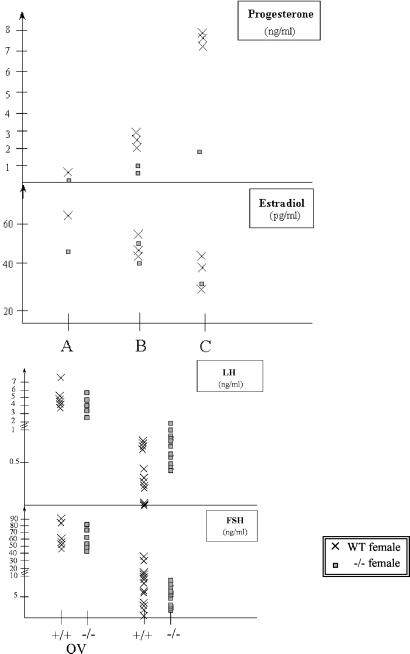
The alpha-fetoprotein gene (Afp) is a member of a multigenic family that comprises the related genes encoding albumin, alpha-albumin, and vitamin D binding protein. The biological role of this major embryonic serum protein is unknown although numerous speculations have been made. We have used gene targeting to show that AFP is not required for embryonic development. AFP null embryos develop normally, and individually transplanted homozygous embryos can develop in an AFP-deficient microenvironment. Whereas mutant homozygous adult males are viable and fertile, AFP null females are infertile. Our analyses of these mice indicate that the defect is caused by a dysfunction of the hypothalamic/pituitary system, leading to anovulation.
Figures
References
-
- Andrews G K, Dziadek M, Tamaoki T. J Biol Chem. 1982;257:5148–5153. - PubMed
-
- Leighton P C, Kitau M J, Chard T, Gordon Y B, Leek A E. Lancet. 1975;7943:1012–1015. - PubMed
-
- Cuckle H S, Wald N J, Lindenbaum R H. Lancet. 1984;8383:926–929. - PubMed
-
- Tilghman S M. In: Oxford Survey on Eukaryotic Genes. Maclean N, editor. Oxford: Oxford Univ. Press; 1985. pp. 160–206. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
