

Two distinct phosphoinositide 3-kinases mediate polypeptide growth factor-stimulated PKB activation
- PMID: 12356726
- PMCID: PMC129034
- DOI: 10.1093/emboj/cdf512
Two distinct phosphoinositide 3-kinases mediate polypeptide growth factor-stimulated PKB activation
Abstract
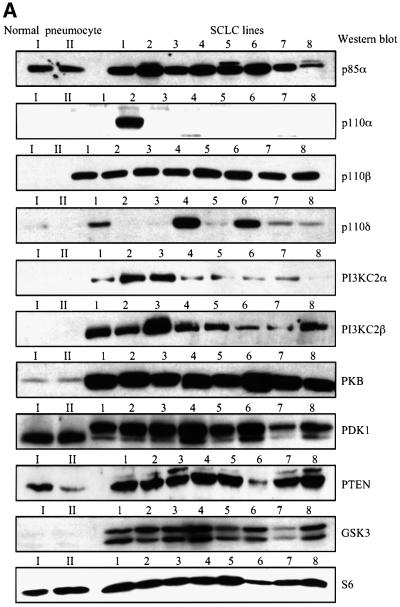
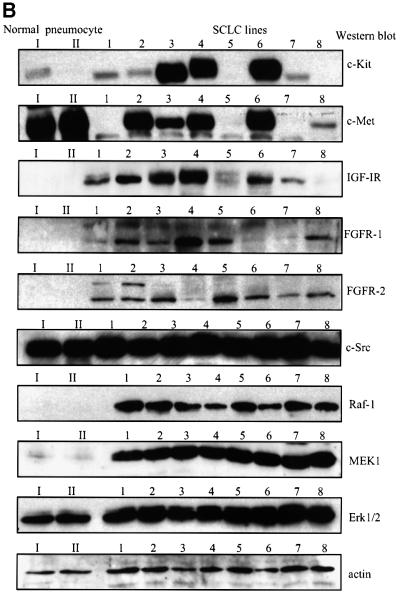
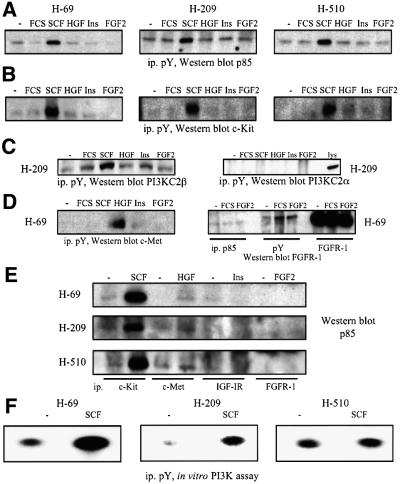
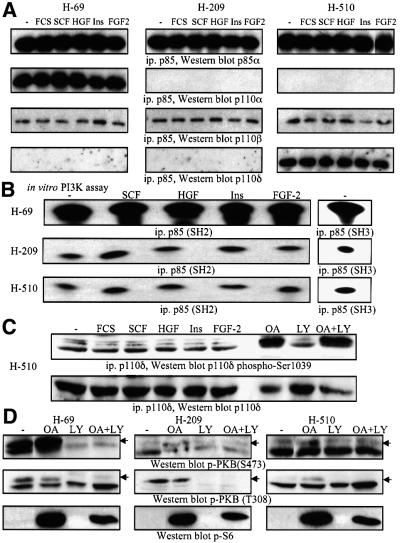
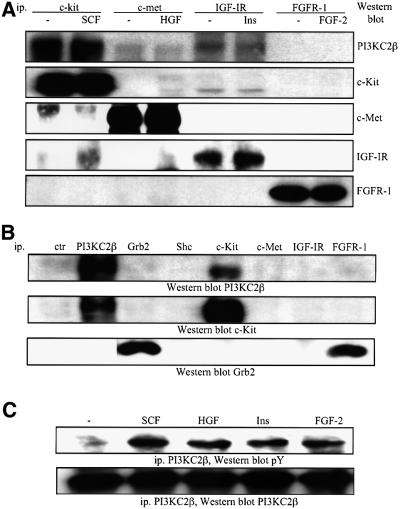
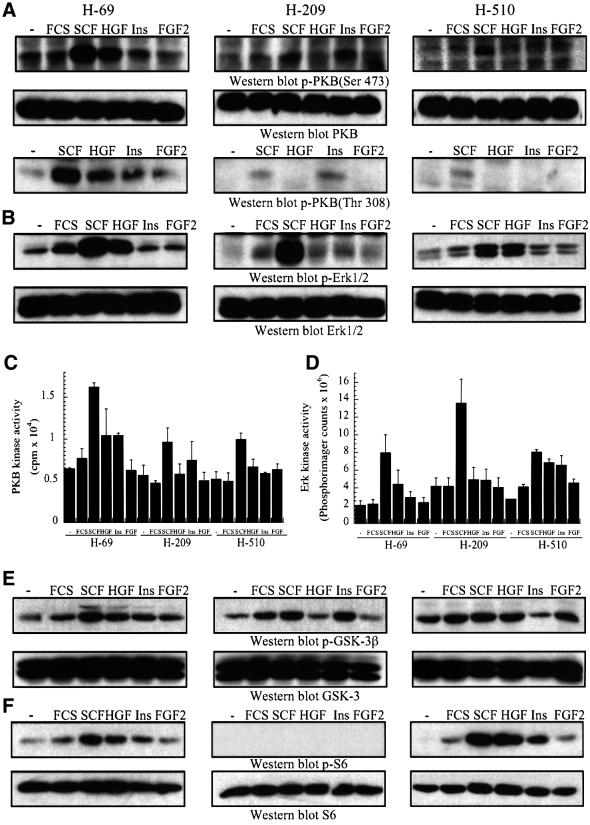
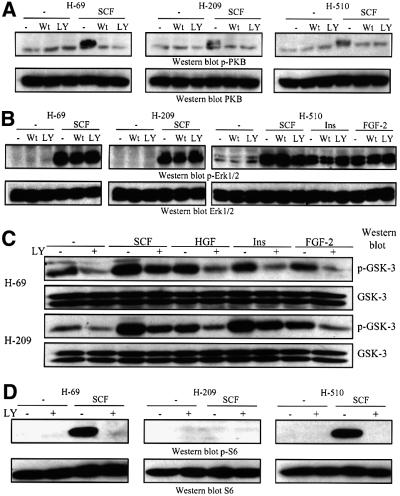
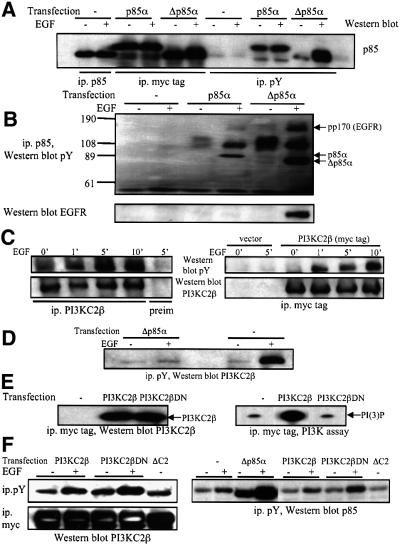
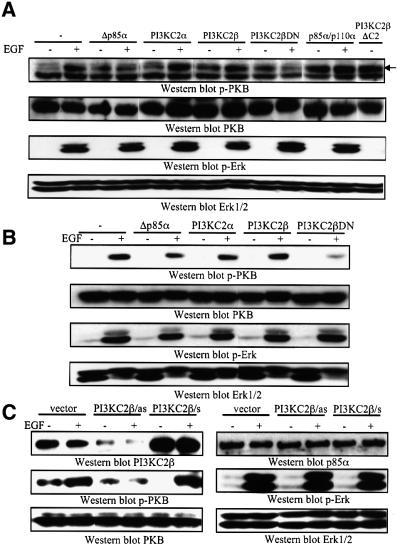
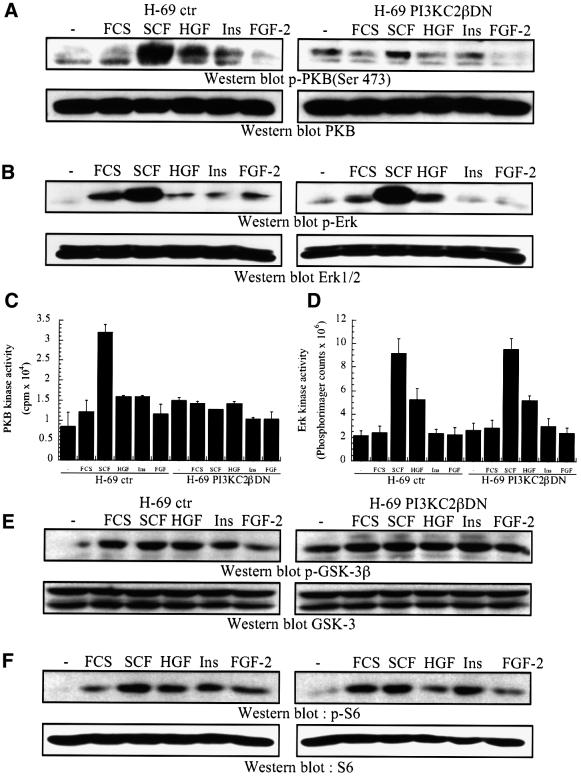
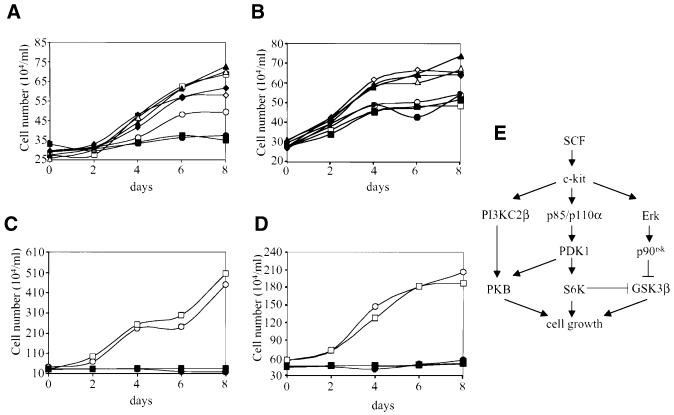
Eight human isoforms of phosphoinositide 3-kinases (PI3Ks) exist, but their individual functions remain poorly understood. Here, we show that different human small cell lung carcinoma (SCLC) cell lines overexpress distinct subsets of class I(A) and II PI3Ks, which results in striking differences in the signalling cascades activated by stem cell factor (SCF). Over expression of class I(A) p85/p110alpha in SCLC cells increased SCF-stimulated protein kinase B (PKB) activation and cell growth, but did not affect extracellular signal-regulated kinase (Erk) or glycogen synthase kinase-3 (GSK-3). This effect was selective, since it was not observed in SCLC cell lines overexpressing p85/p110beta or p85/p110delta. The SCF receptor associated with both class I(A) p85 and class II PI3KC2beta, and both enzymes contributed to SCF-stimulated PKB activity. A dominant-negative PI3KC2beta blocked both PKB activation and SCLC cell growth in response to SCF. Together our data provide novel insights into the specificity and functional significance of PI3K signalling in human cancer.
Figures
References
-
- Alessi D.R., James,S.R., Downes,C.P., Holmes,A.B., Gaffney,P., Reece,C. and Cohen,P. (1997) Purification and characterization of a phosphatidylinositol 3,4,5 trisphosphate-dependent protein kinase (PDK1) which phosphorylates and activates protein kinase Bα. Curr. Biol., 7, 261–269. - PubMed
-
- Alessi D.R., Kozlowski,M.T., Weng,Q.P., Morrice,N. and Avruch,J. (1998) 3-Phosphoinositide-dependent protein kinase 1 (PDK1) phosphorylates and activates the p70 S6 kinase in vivo and in vitro. Curr. Biol., 8, 69–81. - PubMed
-
- Arcaro A., Volinia,S., Zvelebil,M.J., Stein,R., Watton,S.J., Layton,M.J., Gout, I., Ahmadi, K., Downward,J. and Waterfield,M.D. (1998) Human phosphoinositide 3-kinase C2β, the role of calcium and the C2 domain in enzyme activity. J. Biol. Chem., 273, 33082–33090. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
