

Monitoring intermediate folding states of the td group I intron in vivo
- PMID: 12356744
- PMCID: PMC129027
- DOI: 10.1093/emboj/cdf504
Monitoring intermediate folding states of the td group I intron in vivo
Abstract
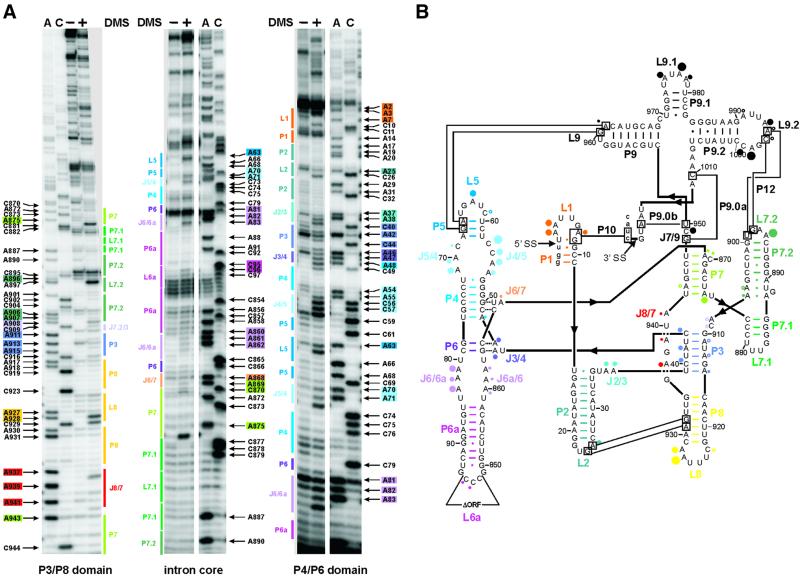
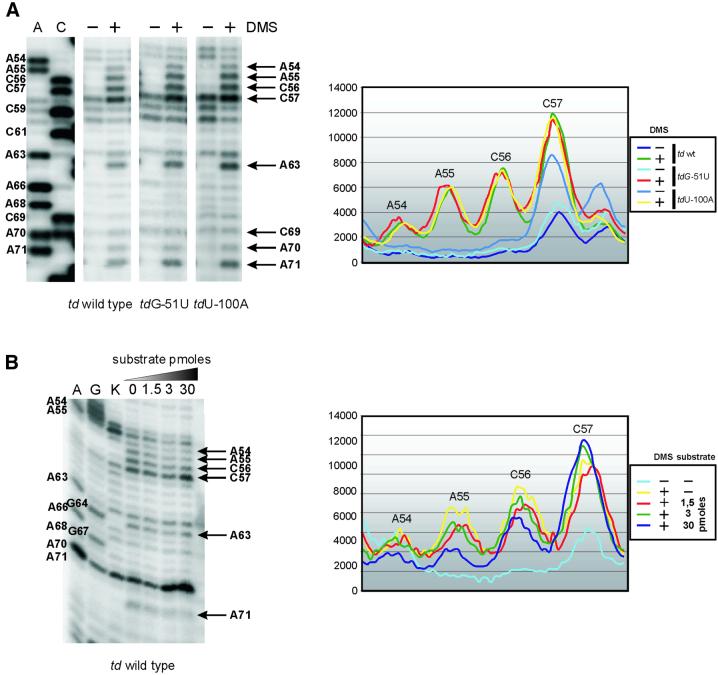
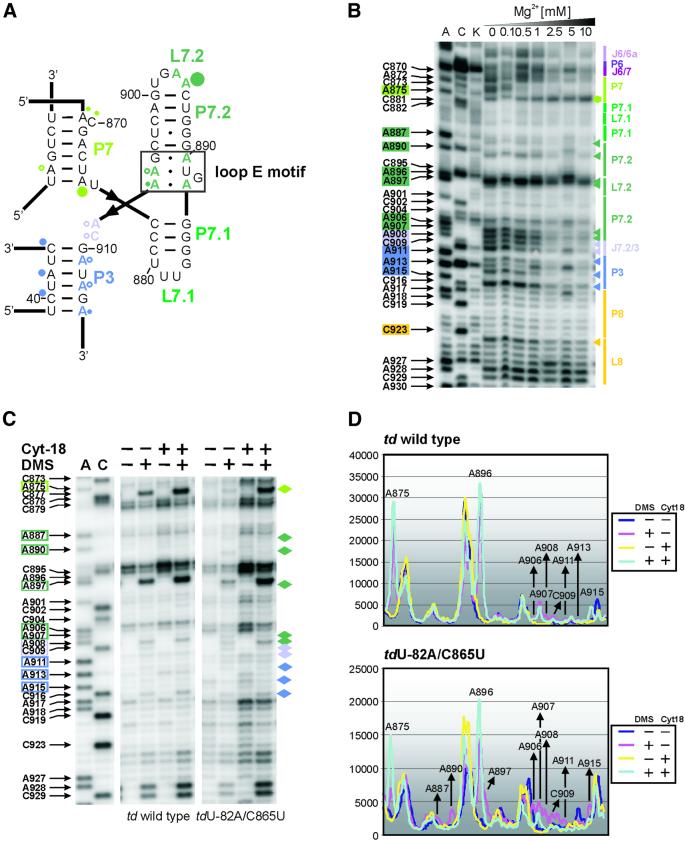
Group I introns consist of two major structural domains, the P4-P6 and P3-P9 domains, which assemble through interactions with peripheral extensions to fold into an active ribozyme. To assess group I intron folding in vivo, we probed the structure of td wild-type and mutant introns using dimethyl sulfate. The results suggest that the majority of the intron population is in the native state in accordance with the current structural model, which was refined to include two novel tertiary contacts. The importance of the loop E motif in the P7.1-P7.2 extension in assisting ribozyme folding was deduced from modeling and mutational analyses. Destabilization of stem P6 results in a deficiency in tertiary structure formation in both major domains, while weakening of stem P7 only interferes with folding of the P3-P9 domain. The different impact of mutations on the tertiary structure suggests that they interfere with folding at different stages. These results provide a first insight into the structure of folding intermediates and suggest a putative order of events in a hierarchical folding pathway in vivo.
Figures


References
-
- Ban N., Nissen,P., Hansen,J., Moore,P.B. and Steitz,T.A. (2000) The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science, 289, 905–920. - PubMed
-
- Belfort M., Chandry,P.S. and Pedersen-Lane,J. (1987) Genetic delineation of functional components of the group I intron in the phage T4 td gene. Cold Spring Harb. Symp. Quant. Biol., 52, 181–192. - PubMed
-
- Caprara M.G., Lehnert,V., Lambowitz,A.M. and Westhof,E. (1996a) A tyrosyl-tRNA synthetase recognizes a conserved tRNA-like structural motif in the group I intron catalytic core. Cell, 87, 1135–1145. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
