

T cell receptor ligation induces the formation of dynamically regulated signaling assemblies
- PMID: 12356870
- PMCID: PMC2173229
- DOI: 10.1083/jcb.200203043
T cell receptor ligation induces the formation of dynamically regulated signaling assemblies
Abstract
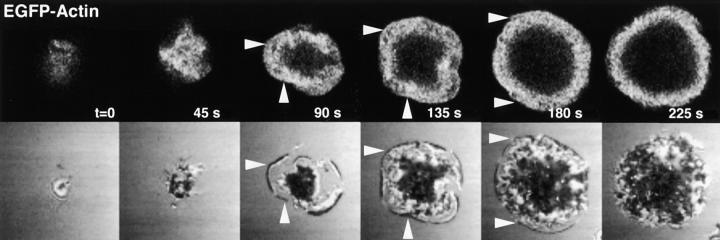
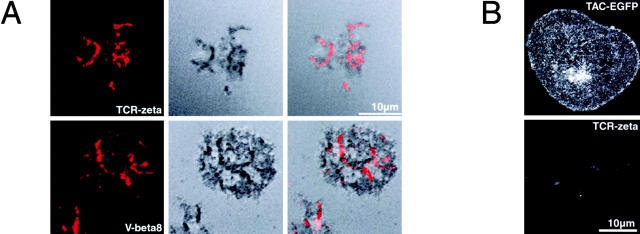
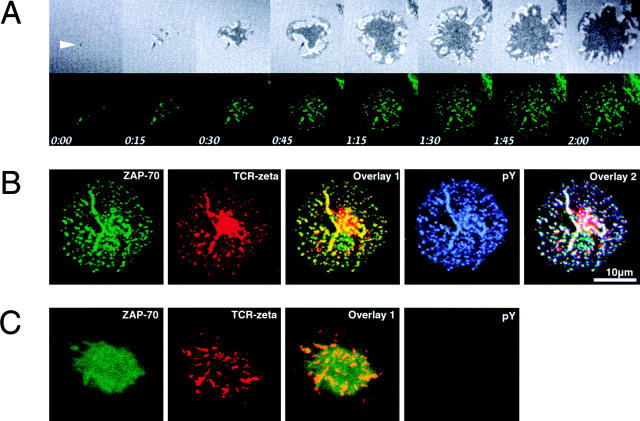
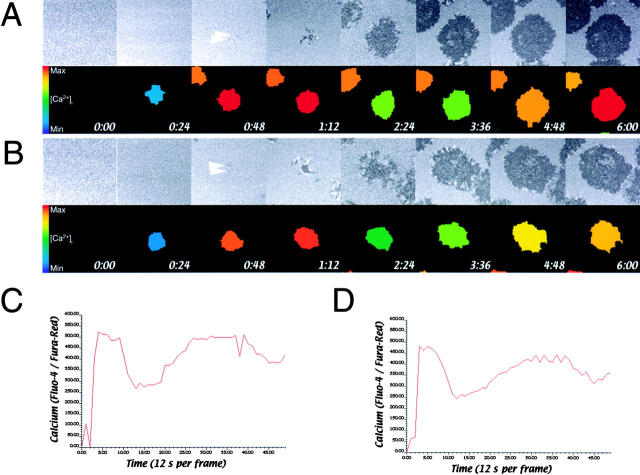
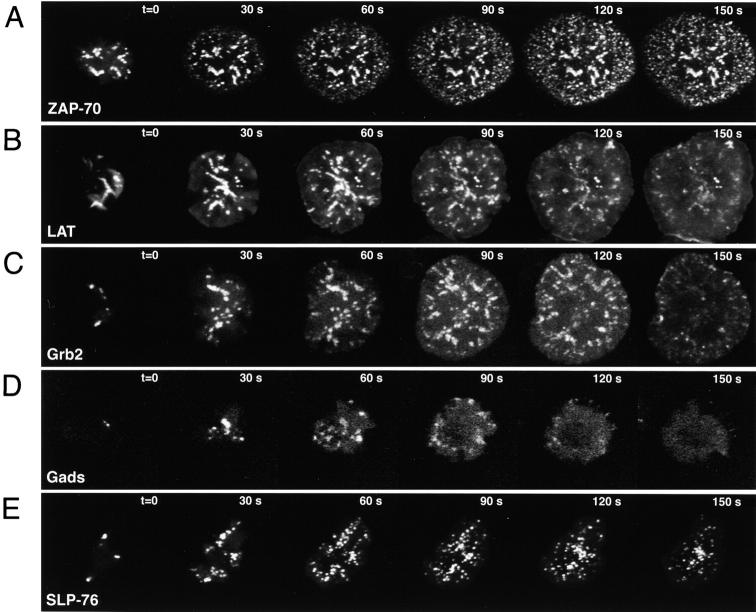
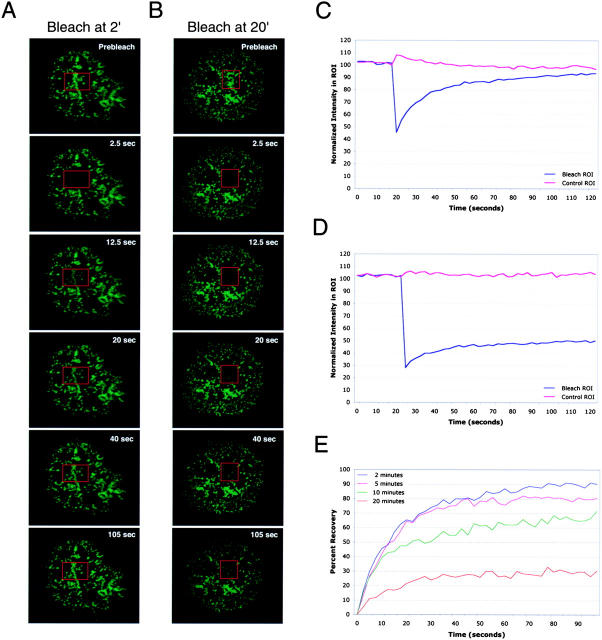
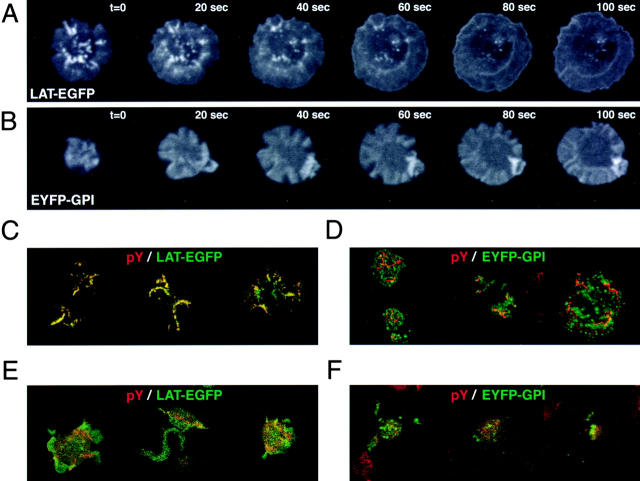
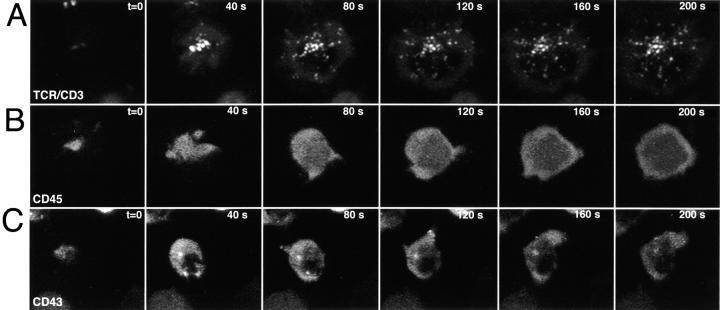
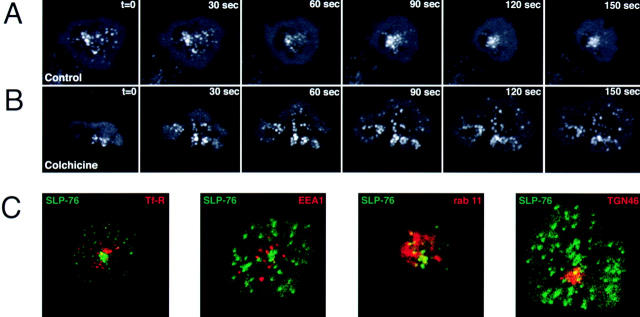
Tcell antigen receptor (TCR) ligation initiates tyrosine kinase activation, signaling complex assembly, and immune synapse formation. Here, we studied the kinetics and mechanics of signaling complex formation in live Jurkat leukemic T cells using signaling proteins fluorescently tagged with variants of enhanced GFP (EGFP). Within seconds of contacting coverslips coated with stimulatory antibodies, T cells developed small, dynamically regulated clusters which were enriched in the TCR, phosphotyrosine, ZAP-70, LAT, Grb2, Gads, and SLP-76, excluded the lipid raft marker enhanced yellow fluorescent protein-GPI, and were competent to induce calcium elevations. LAT, Grb2, and Gads were transiently associated with the TCR. Although ZAP-70-containing clusters persisted for more than 20 min, photobleaching studies revealed that ZAP-70 continuously dissociated from and returned to these complexes. Strikingly, SLP-76 translocated to a perinuclear structure after clustering with the TCR. Our results emphasize the dynamically changing composition of signaling complexes and indicate that these complexes can form within seconds of TCR engagement, in the absence of either lipid raft aggregation or the formation of a central TCR-rich cluster.
Figures

References
-
- Allenspach, E.J., P. Cullinan, J. Tong, Q. Tang, A.G. Tesciuba, J.L. Cannon, S.M. Takahashi, R. Morgan, J.K. Burkhardt, and A.I. Sperling. 2001. ERM-dependent movement of CD43 defines a novel protein complex distal to the immunological synapse. Immunity. 15:739–750. - PubMed
-
- Anton van der Merwe, P., S.J. Davis, A.S. Shaw, and M.L. Dustin. 2000. Cytoskeletal polarization and redistribution of cell-surface molecules during T cell antigen recognition. Semin. Immunol. 12:5–21. - PubMed
-
- Balamuth, F., D. Leitenberg, J. Unternaehrer, I. Mellman, and K. Bottomly. 2001. Distinct patterns of membrane microdomain partitioning in Th1 and Th2 cells. Immunity. 15:729–738. - PubMed
-
- Boniface, J.J., J.D. Rabinowitz, C. Wulfing, J. Hampl, Z. Reich, J.D. Altman, R.M. Kantor, C. Beeson, H.M. McConnell, and M.M. Davis. 1998. Initiation of signal transduction through the T cell receptor requires the multivalent engagement of peptide/MHC ligands. Immunity. 9:459–466. - PubMed
-
- Brdicka, T., D. Pavlistova, A. Leo, E. Bruyns, V. Korinek, P. Angelisova, J. Scherer, A. Shevchenko, I. Hilgert, J. Cerny, et al. 2000. Phosphoprotein associated with glycosphingolipid-enriched microdomains (PAG), a novel ubiquitously expressed transmembrane adaptor protein, binds the protein tyrosine kinase csk and is involved in regulation of T cell activation. J. Exp. Med. 191:1591–1604. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
