

Presynaptic internal Ca2+ stores contribute to inhibitory neurotransmitter release onto mouse cerebellar Purkinje cells
- PMID: 12359635
- PMCID: PMC1573523
- DOI: 10.1038/sj.bjp.0704901
Presynaptic internal Ca2+ stores contribute to inhibitory neurotransmitter release onto mouse cerebellar Purkinje cells
Abstract
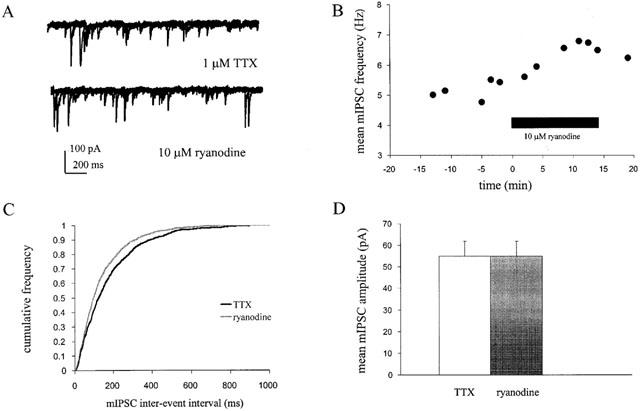
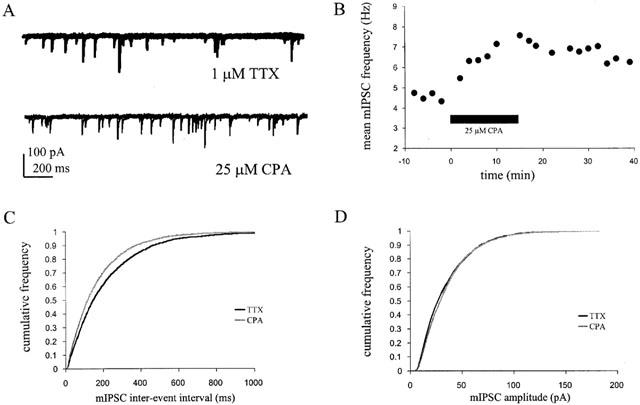
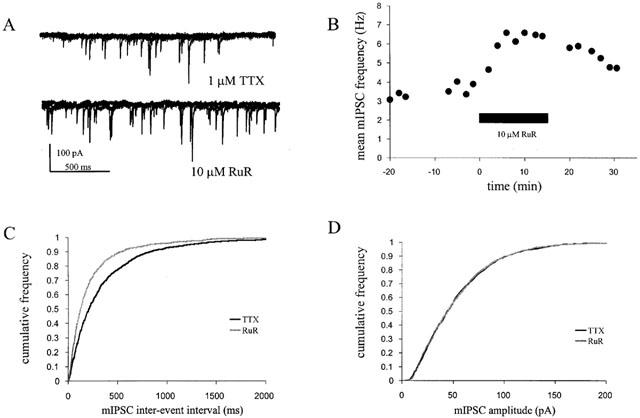
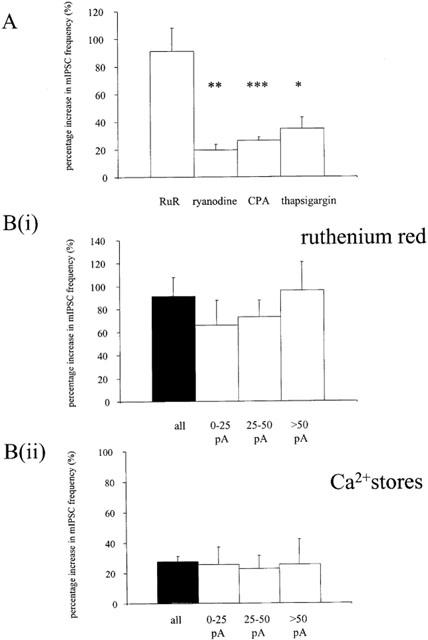
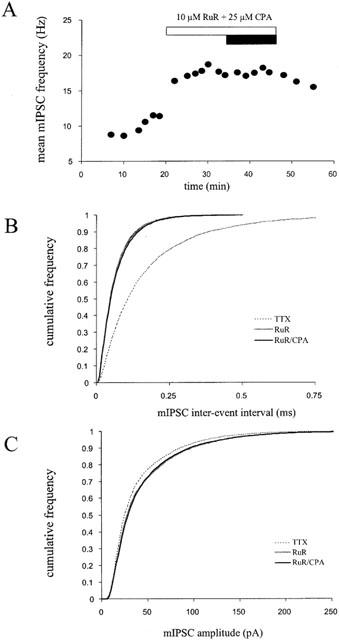
1. Miniature inhibitory postsynaptic currents (mIPSCs) were recorded in mouse Purkinje cells in the presence of 1 micro M tetrodotoxin (TTX). Under these conditions, which eliminated Ca(2+) influx through voltage-dependent Ca(2+) channels (VDCCs), the contribution of Ca(2+) stores to spontaneous GABA release was examined. 2. The plant alkaloid ryanodine acts as an inhibitor of endoplasmic reticulum ryanodine-sensitive Ca(2+) release channels (ryanodine receptors) at low micromolar concentrations. Ryanodine effects were confined to a subpopulation of cells tested. At 10 micro M ryanodine, 4/12 cells showed a significant increase in mean mIPSC frequency of +19.6+/-4.0% (n=4). 3. The sarco/endoplasmic reticulum Ca(2+)-ATPase (SERCA) pump inhibitor cyclopiazonic acid (CPA) produced a more robust effect. In 8/10 cells, 25 micro M CPA caused a significant increase in mean mIPSC frequency; the mean increase being +26.0+/-3.0% (n=8). Similar results were seen with thapsigargin (1-2 micro M), another SERCA pump inhibitor. 4. Ruthenium red (RuR) has been proposed to either act directly on the release machinery or block Ca(2+) pumps on internal stores. At 10 micro M RuR, all cells showed a rapid, large increase in mean mIPSC frequency of +90.4+/-16.4% (n=9). This increase was greater than that seen by agents known to modulate Ca(2+) stores and was more consistent with a direct action. At this concentration, RuR also occluded the effects of CPA. 5. For all reagents, there were no obvious effects on mean mIPSC amplitude. However, the effects on mIPSC frequency were consistent with a presynaptic action and indicate that Ca(2+) stores may contribute to spontaneous GABA release onto mouse Purkinje cells.
Figures
Similar articles
-
Glycinergic mIPSCs in mouse and rat brainstem auditory nuclei: modulation by ruthenium red and the role of calcium stores.J Physiol. 2003 Feb 1;546(Pt 3):691-9. doi: 10.1113/jphysiol.2002.035071. J Physiol. 2003. PMID: 12562997 Free PMC article.
-
Calcium release from presynaptic internal stores is required for ethanol to increase spontaneous gamma-aminobutyric acid release onto cerebellum Purkinje neurons.J Pharmacol Exp Ther. 2007 Oct;323(1):356-64. doi: 10.1124/jpet.107.126144. Epub 2007 Jul 25. J Pharmacol Exp Ther. 2007. PMID: 17652632
-
Mechanism of GABA receptor-mediated inhibition of spontaneous GABA release onto cerebellar Purkinje cells.Eur J Neurosci. 2004 Aug;20(3):684-700. doi: 10.1111/j.1460-9568.2004.03505.x. Eur J Neurosci. 2004. PMID: 15255979
-
Presynaptic inhibition of elicited neurotransmitter release.Trends Neurosci. 1997 May;20(5):204-12. doi: 10.1016/s0166-2236(96)01015-6. Trends Neurosci. 1997. PMID: 9141196 Review.
-
Presynaptic calcium stores and synaptic transmission.Curr Opin Neurobiol. 2005 Jun;15(3):275-81. doi: 10.1016/j.conb.2005.05.003. Curr Opin Neurobiol. 2005. PMID: 15919193 Review.
Cited by
-
The phytocannabinoid Delta(9)-tetrahydrocannabivarin modulates inhibitory neurotransmission in the cerebellum.Br J Pharmacol. 2008 May;154(1):204-15. doi: 10.1038/bjp.2008.57. Epub 2008 Mar 3. Br J Pharmacol. 2008. PMID: 18311186 Free PMC article.
-
Presynaptic ryanodine receptors are required for normal quantal size at the Caenorhabditis elegans neuromuscular junction.J Neurosci. 2005 Jul 20;25(29):6745-54. doi: 10.1523/JNEUROSCI.1730-05.2005. J Neurosci. 2005. PMID: 16033884 Free PMC article.
-
Ethanol-enhanced GABA release: a focus on G protein-coupled receptors.Brain Res Rev. 2011 Jan 1;65(2):113-23. doi: 10.1016/j.brainresrev.2010.09.003. Epub 2010 Sep 15. Brain Res Rev. 2011. PMID: 20837058 Free PMC article. Review.
-
Action potential-evoked and ryanodine-sensitive spontaneous Ca2+ transients at the presynaptic terminal of a developing CNS inhibitory synapse.J Neurosci. 2004 Aug 4;24(31):6946-57. doi: 10.1523/JNEUROSCI.1397-04.2004. J Neurosci. 2004. PMID: 15295030 Free PMC article.
-
Roles and Sources of Calcium in Synaptic Exocytosis.Adv Neurobiol. 2023;33:139-170. doi: 10.1007/978-3-031-34229-5_6. Adv Neurobiol. 2023. PMID: 37615866
References
-
- AUGUSTINE G.J. How does calcium trigger neurotransmitter release. Curr. Opin. Neurobiol. 2001;11:320–326. - PubMed
-
- BERRIDGE M.J. Neuronal calcium signaling. Neuron. 1998;21:13–26. - PubMed
-
- BEUTNER G., SHARMA V.K., GIOVANNUCCI D.R., YULE D.I., SHEU S.S. Identification of a ryanodine receptor in rat heart mitochondria. J. Biol. Chem. 2001;276:21482–21488. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
