

In the presence of L-NAME SERCA blockade induces endothelium-dependent contraction of mouse aorta through activation of smooth muscle prostaglandin H2/thromboxane A2 receptors
- PMID: 12359637
- PMCID: PMC1573510
- DOI: 10.1038/sj.bjp.0704884
In the presence of L-NAME SERCA blockade induces endothelium-dependent contraction of mouse aorta through activation of smooth muscle prostaglandin H2/thromboxane A2 receptors
Abstract
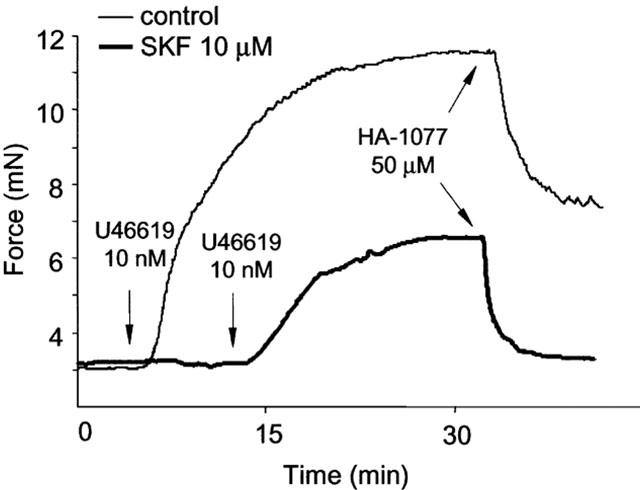
1. The mechanism of transient contractions induced by the sarcoplasmic-endoplasmic reticulum calcium ATPase (SERCA) blocker cyclopiazonic acid (CPA) in the presence of L-NAME was investigated in mouse aorta. 2. The contractions elicited by 10 micro M CPA required an intact endothelium, were dependent upon external Ca(2+) and were prevented by 10 micro M indomethacin, the inhibitor of prostaglandin synthesis, or 1 micro M SQ29548, the specific prostaglandin H2/thromboxane A2 (PGH2/TXA2) receptor blocker. 3. A blocker of receptor/store operated Ca(2+) channels and voltage gated calcium channels (VGCC), SK&F 96365 (10 micro M), completely abolished the contractions, while a specific blocker of VGCC nifedipine (1 micro M) inhibited them by one third. 4. Dichlorobenzamyl hydrochloride, a blocker of Na(+)/Ca(2+) exchange effectively prevented return of tension to baseline value. 5. At higher concentrations (30-100 micro M) CPA induced indomethacin-resistant tonic contractions of mouse aorta. The CPA dose response curve for tonic contractions is shifted to the right compared to the transient contractions suggesting that smooth muscle is less sensitive to CPA than endothelium. 6. PGH2/TXA2 receptors in mouse aorta are highly sensitive to the thromboxane analogue U46619 (EC(50) : 1.93 nM). This compound stimulates contractions even in the absence of external Ca(2+), which are abolished by the Rho-kinase inhibitor HA-1077. 7. The results suggest that 10 micro M CPA induced capacitive Ca(2+) entry in endothelial cells stimulating the release of PGH2/TXA2, which subsequently caused smooth muscle contraction dependent on Ca(2+) influx and myofilament sensitization by Rho-kinase. Higher concentrations of CPA (30-100 micro M) directly induced contraction of mouse aortic smooth muscle.
Figures









References
-
- AUCH-SCHWELK W., KATUSIC Z.C., VANHOUTTE P.M. Thromboxane A2 receptor antagonists inhibit endothelium-dependent contractions. Hypertension. 1990;15:699–703. - PubMed
-
- BLAUSTEIN M.P., LEDERER W.J. Sodium/calcium exchange: its phisiological implication. Physiological reviews. 1999;79:763–854. - PubMed
-
- CHANNON J.Y., LESLIE C.C. A calcium-dependent mechanism for associating a soluble arachidonoyl-hydrolyzing phospholipase A2 with membrane in the macrophage cell line RAW 264.7. J. Biol. Chem. 1990;265:5409–5413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
