

The Ccz1-Mon1 protein complex is required for the late step of multiple vacuole delivery pathways
- PMID: 12364329
- PMCID: PMC2754690
- DOI: 10.1074/jbc.M208191200
The Ccz1-Mon1 protein complex is required for the late step of multiple vacuole delivery pathways
Abstract
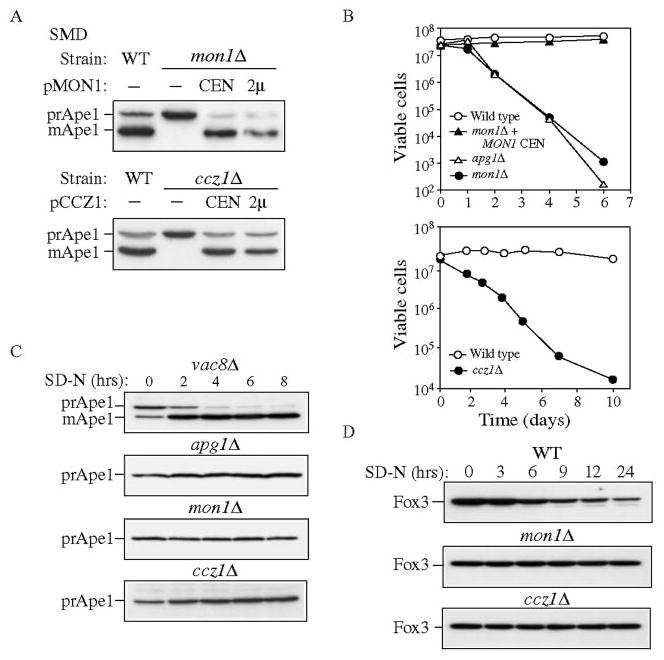
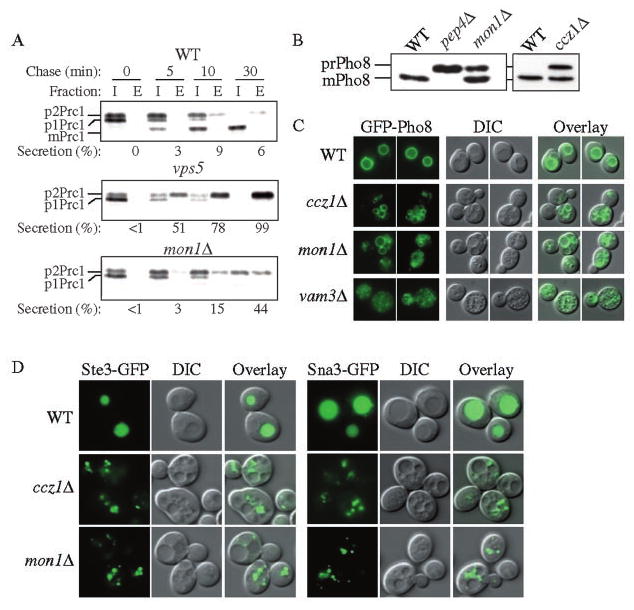
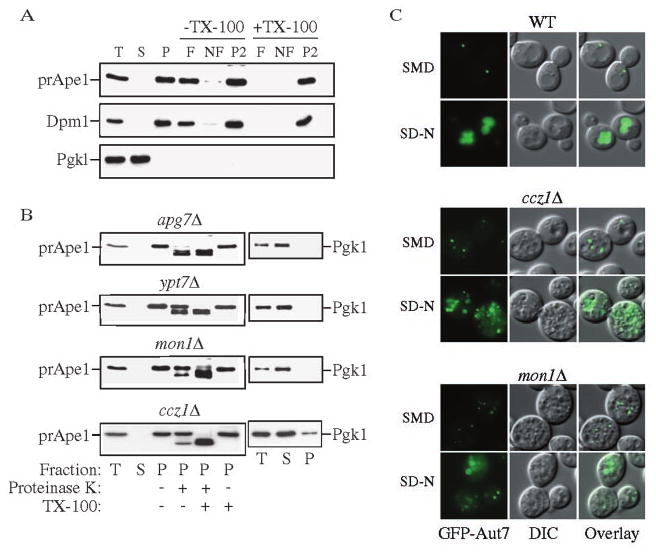
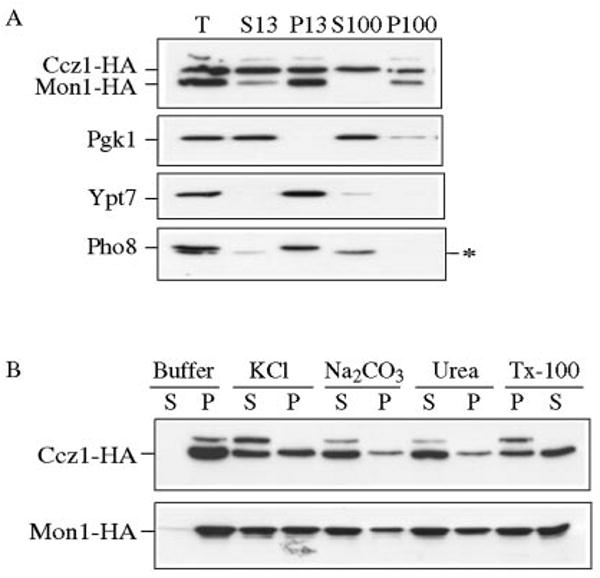
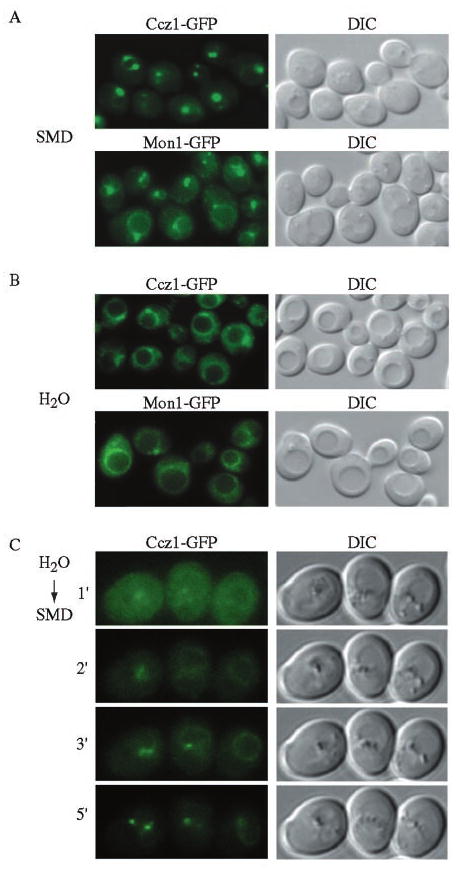
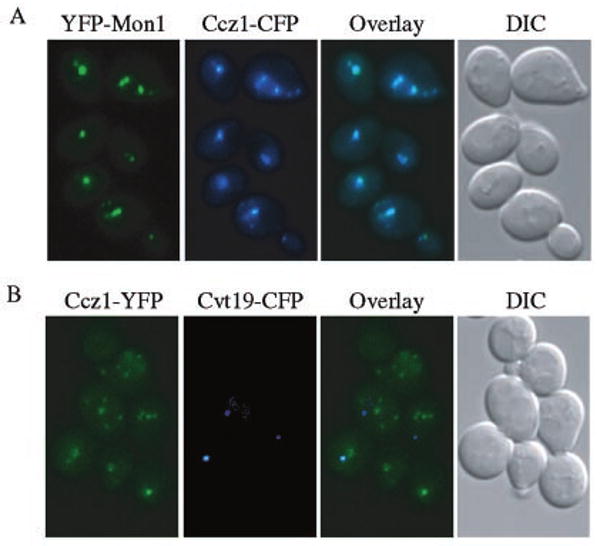
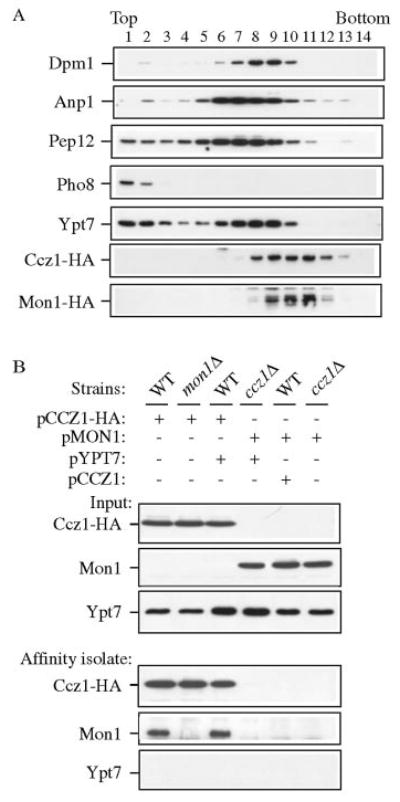
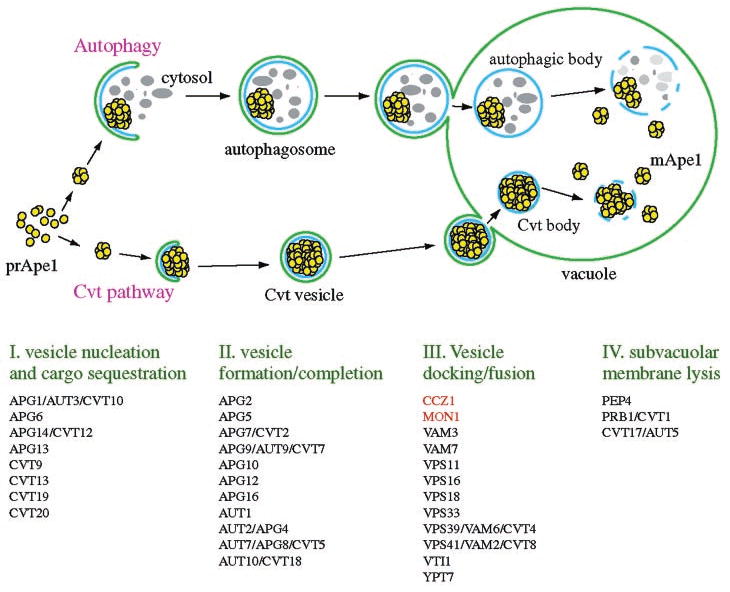
Mon1 and Ccz1 were identified from a gene deletion library as mutants defective in the vacuolar import of aminopeptidase I (Ape1) via the cytoplasm to vacuole targeting (Cvt) pathway. The mon1Delta and ccz1Delta strains also displayed defects in autophagy and pexophagy, degradative pathways that share protein machinery and mechanistic features with the biosynthetic Cvt pathway. Further analyses indicated that Mon1, like Ccz1, was required in nearly all membrane-trafficking pathways where the vacuole represented the terminal acceptor compartment. Accordingly, both deletion strains had kinetic defects in the biosynthetic delivery of resident vacuolar hydrolases through the CPY, ALP, and MVB pathways. Biochemical and microscopy studies suggested that Mon1 and Ccz1 functioned after transport vesicle formation but before (or at) the fusion step with the vacuole. Thus, ccz1Delta and mon1Delta are the first mutants identified in screens for the Cvt and Apg pathways that accumulate precursor Ape1 within completed cytosolic vesicles. Subcellular fractionation and co-immunoprecipitation experiments confirm that Mon1 and Ccz1 physically interact as a stable protein complex termed the Ccz1-Mon1 complex. Microscopy of Ccz1 and Mon1 tagged with a fluorescent marker indicated that the Ccz1-Mon1 complex peripherally associated with a perivacuolar compartment and may attach to the vacuole membrane in agreement with their proposed function in fusion.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
