

doi: 10.1101/gad.240002.
Bicoid associates with the 5'-cap-bound complex of caudal mRNA and represses translation
Affiliations
- PMID: 12368268
- PMCID: PMC187448
- DOI: 10.1101/gad.240002
Item in Clipboard
Bicoid associates with the 5'-cap-bound complex of caudal mRNA and represses translation
Genes Dev.
.
Abstract
Translational control plays a key role in many biological processes including pattern formation during early Drosophila embryogenesis. In this process, the anterior determinant Bicoid (BCD) acts not only as a transcriptional activator of segmentation genes but also causes specific translational repression of ubiquitously distributed caudal (cad) mRNA in the anterior region of the embryo. We show that translational repression of cad mRNA is dependent on a functional eIF4E-binding motif. The results suggest a novel mode of translational repression, which combines the strategy of target-specific binding to 3'-untranslated sequences and interference with 5'-cap-dependent translation initiation in one protein.
Figures
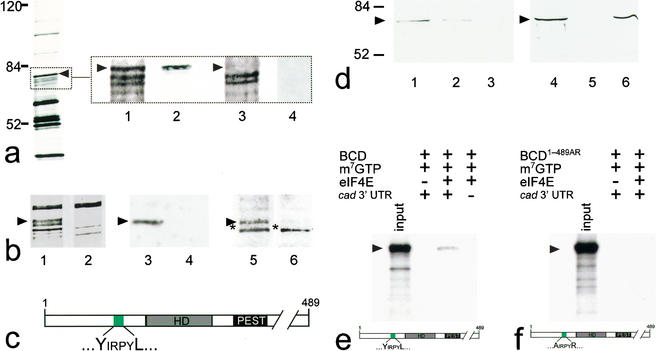
BCD copurifies with 5′-cap-bound proteins. Cytoplasmic protein extracts of young embryos were affinity-purified using a cap-analog m7GTP-sepharose resin (Edery et al. 1988). (a) Silver-stained SDS-PAGE of affinity-purified proteins contained within cytoplasmic extracts of wild-type and bcd mutant Drosophila embryos. Arrowhead marks the protein band that was subsequently identified as BCD (left, see also lanes 1–4 in b). Note that eIF4E, the most abundant 30-kD component among the purified proteins, is run off the gel (left) to obtain maximum resolution of the relevant range of protein bands between 50 and 100 kD. (Lanes 1–4) Relevant portions of silver-stained SDS-PAGE (dotted box) including a ∼70-kD protein (lane 1, arrowhead) that is recognized by monospecific anti-BCD antibodies on Western blots (lane 2). This protein is not present in silver-stained SDS-PAGE (lane 3), and anti-BCD antibodies stained Western blots (lane 4) of corresponding extracts from embryos that derived from homozygous bcd mutant females. (b) Relevant portions of silver-stained SDS-PAGE showing that the GFP–BCD fusion protein (lane 1, 95 kD) is absent in wild-type embryos (lane 2). Corresponding Western blots show that the 95-kD fusion protein reacts with both anti-BCD (lane 3; lane 4 is a wild-type control lacking the GFP–BCD transgene) and anti-GFP antibodies (lane 5; lane 6 is a wild-type control lacking the GFP–BCD transgene; asterisk marks cross-reacting protein of unknown identity). (c) Schematic diagram of BCD (positions refer to sequence according to Berleth et al. 1988) showing the homeodomain (HD, gray box; position 91–154), the PEST domain (PEST, black box; position 170–203), and a YIRPYL motif (green box; position 68–73). The eIF4E-binding properties of this motif have been recently analyzed in great detail in the context of human BP1 (Marcotrigiano et al. 1999; for review, see Raught et al. 2000; Sachs and Varani 2000; Miron et al. 2001). (d) Western blots showing that BCD bound to m7GTP-sepharose coupled recombinant eIF4E (lanes 1,4) can be competed for with 100 nM (lane 2) and 1 mM (lanes 3,5) of a peptide containing the YDRKFL motif of human BP1, whereas 1 mM of a mutated human BP1 peptide (mutated motif is ADRKFR) did not interfere with the binding of BCD to eIF4E (lane 6). (e,f) Autoradiogram showing that in vitro translated 35S-labeled BCD (input; a schematic representation of the protein is shown at the bottom of e) is capable of interacting with m7GTP-sepharose-bound recombinant eIF4E in the presence of in vitro transcribed cad 3′-UTR (e), whereas in vitro translated 35S-labeled BCD1–489AR (input; a schematic representation of the protein is shown at the bottom of f) showed no significant binding to eIF4E (f). Note that there is no unspecific binding of in vitro translated protein to m7GTP-beads without precoupled eIF4E. For details, see Materials and Methods.
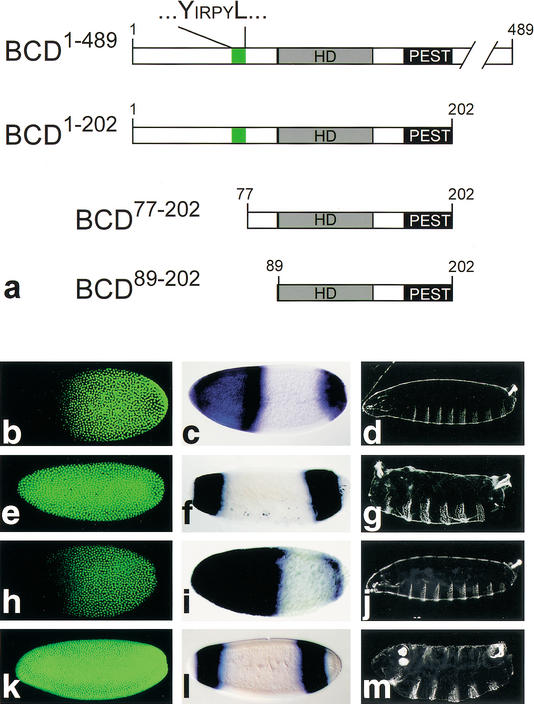
Functional analysis of mutant BCD by transgene-dependent expression in bcd mutant embryos. (a) Schematic representation of the BCD-deletion mutants (cf. Fig. 1c, wild-type BCD). (b–d) Wild-type embryos are characterized by translational repression of cad mRNA (absence of anti-CAD antibody staining (green) in the anterior region of preblastoderm embryos (b; Dubnau and Struhl 1996; Rivera-Pomar et al. 1996), by activation of zygotic hb transcription as revealed by whole mount in situ hybridization with an hb cDNA probe (c; Klingler and Gergen 1993; reviewed in Martinez Arias 1993), and by the wild-type cuticle pattern (d). (e–g) Embryos derived from homozygous bcdE1 mutant females fail to repress cad mRNA translation (e), lack the anterior hb expression domain (which is replaced by a duplication of the posterior, BCD-independent hb expression domain; f), and show a bcd mutant cuticle phenotype (g; Frohnhöfer and Nüsslein-Volhard 1986). (h–j) Transgene-derived BCD1–489 expression restores all aspects of the bcd mutant phenotype including translational repression of cad mRNA (h), anterior hb expression (i), and the larval cuticle phenotype (j). (k–m) cad mRNA translation is not repressed in response to transgene-derived BCD77–202 (k) or BCD89–202 (data not shown). In both cases, the transgene-expressed BCD deletion mutants also fail to restore anterior hb expression (l) and develop a bcd mutant cuticle phenotype (m) because of the absence of the C-terminal transactivation domains (Sauer et al. 1995; Schaeffer et al. 1999). Orientation of embryos is anterior left and dorsal up.
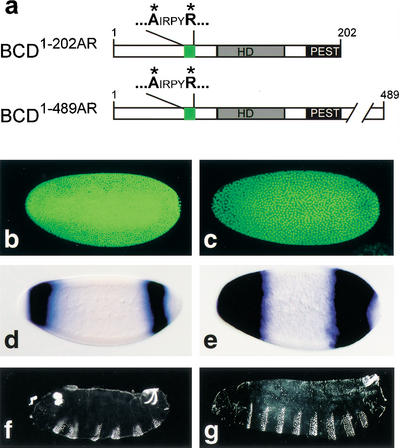
Translational repression of cad mRNA, transcriptional activation of anterior hb expression, and rescue of segmentation defects in response to transgene-expressed BCD replacement mutants. (a) Schematic representation of the BCD replacement mutants BCD1–202AR and BCD1–489AR in which the YIRPYL motif is changed into AIRPYR (green box; for other details, see Fig. 1c). (b,d,f) Embryos from homozygous bcdE1 females expressing transgene-derived BCD1–202AR fail to repress cad mRNA translation (b) or to activate anterior hb expression (d), and develop a bcd mutant cuticle phenotype (f). (c,e,g) Embryos from homozygous bcdE1 females expressing transgene-derived BCD1–489AR fail to repress cad mRNA translation (c) but activate anterior hb expression (e) and develop the normal head and thoracic pattern elements (g). For further details, see text.
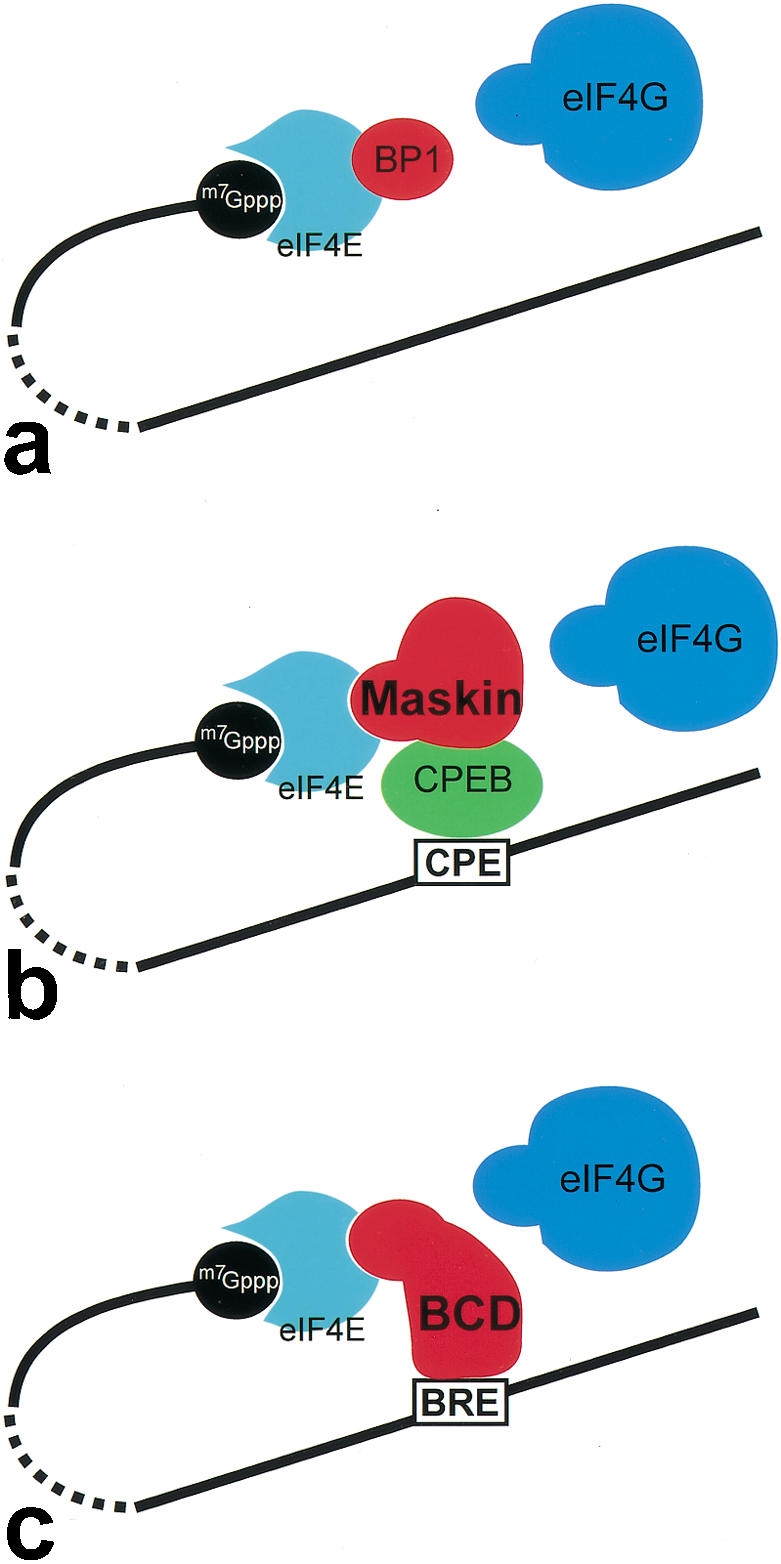
Different modes of cap-dependent translational repression by interference with the assembly of the eIF4E ∷ eIF4G interaction complex. (a) Binding of BP to eIF4E blocks eIF4G-binding and modulates translation efficiency of 5′-capped mRNAs in an insulin signaling-dependent manner (for review, see Raught et al. 2000; Sachs and Varani 2000). (b) Translational repression of mRNAs, which anchor CPEB through a CP element in their 3′-UTR. CPEB is able to associate with Maskin, which successively blocks the eIF4E ∷ eIF4G interaction by binding to eIF4E (for review, see Richter 2000; Richter and Theurkauf 2001). (c) BCD uses a similar strategy of repression by combining the binding properties of both CPEB and Maskin. BCD binds directly to the BRE in the 3′-UTR and blocks the eIF4E ∷ eIF4G interaction at the 5′-end. In each case (a–c), the eIF4E interaction involves the YxxxxL motif of the translational repressors.
Similar articles
-
Sequence interval within the PEST motif of Bicoid is important for translational repression of caudal mRNA in the anterior region of the Drosophila embryo.EMBO J. 1999 Apr 1;18(7):1966-73. doi: 10.1093/emboj/18.7.1966. EMBO J. 1999. PMID: 10202159 Free PMC article.
-
A new paradigm for translational control: inhibition via 5'-3' mRNA tethering by Bicoid and the eIF4E cognate 4EHP.Cell. 2005 May 6;121(3):411-23. doi: 10.1016/j.cell.2005.02.024. Cell. 2005. PMID: 15882623
-
RNA recognition and translational regulation by a homeodomain protein.Nature. 1996 Feb 22;379(6567):694-9. doi: 10.1038/379694a0. Nature. 1996. PMID: 8602214
-
Drosophila development: homeodomains and translational control.Curr Biol. 1996 Jul 1;6(7):773-5. doi: 10.1016/s0960-9822(02)00591-2. Curr Biol. 1996. PMID: 8805294 Review.
-
Contrasting mechanisms of regulating translation of specific Drosophila germline mRNAs at the level of 5'-cap structure binding.Biochem Soc Trans. 2005 Dec;33(Pt 6):1544-6. doi: 10.1042/BST0331544. Biochem Soc Trans. 2005. PMID: 16246166 Review.
Cited by
-
The oncogene eIF4E: using biochemical insights to target cancer.J Interferon Cytokine Res. 2013 May;33(5):227-38. doi: 10.1089/jir.2012.0142. Epub 2013 Mar 8. J Interferon Cytokine Res. 2013. PMID: 23472659 Free PMC article. Review.
-
Control of messenger RNA fate by RNA-binding proteins: an emphasis on mammalian spermatogenesis.J Androl. 2012 May-Jun;33(3):309-37. doi: 10.2164/jandrol.111.014167. Epub 2011 Jul 14. J Androl. 2012. PMID: 21757510 Free PMC article. Review.
-
Shape and function of the Bicoid morphogen gradient in dipteran species with different sized embryos.Dev Biol. 2008 Apr 15;316(2):350-8. doi: 10.1016/j.ydbio.2008.01.039. Epub 2008 Feb 13. Dev Biol. 2008. PMID: 18328473 Free PMC article.
-
Crystal structure of a minimal eIF4E-Cup complex reveals a general mechanism of eIF4E regulation in translational repression.RNA. 2012 Sep;18(9):1624-34. doi: 10.1261/rna.033639.112. Epub 2012 Jul 25. RNA. 2012. PMID: 22832024 Free PMC article.
-
Optogenetic control of the Bicoid morphogen reveals fast and slow modes of gap gene regulation.Cell Rep. 2022 Mar 22;38(12):110543. doi: 10.1016/j.celrep.2022.110543. Cell Rep. 2022. PMID: 35320726 Free PMC article.
References
-
- Curtis D, Lehmann R, Zamore PD. Translational regulation in development. Cell. 1995;81:171–178. - PubMed
-
- Driever W. Maternal control of anterior development in the Drosophila embryo. In: Bate M, Martinez Arias A, editors. The development of Drosophila melanogaster. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1993. pp. 301–324.
-
- Driever W, Nüsslein-Volhard C. The Bicoid protein is a positive regulator of hunchback transcription in the early Drosophila embryo. Nature. 1989;337:138–143. - PubMed
-
- Driever W, Ma J, Nüsslein-Volhard C, Ptashne M. Rescue of bicoid mutant Drosophila embryos by bicoid fusion proteins containing heterologous activating sequences. Nature. 1989;342:149–154. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous