

Poliovirus internal ribosome entry segment structure alterations that specifically affect function in neuronal cells: molecular genetic analysis
- PMID: 12368304
- PMCID: PMC136602
- DOI: 10.1128/jvi.76.21.10617-10626.2002
Poliovirus internal ribosome entry segment structure alterations that specifically affect function in neuronal cells: molecular genetic analysis
Abstract
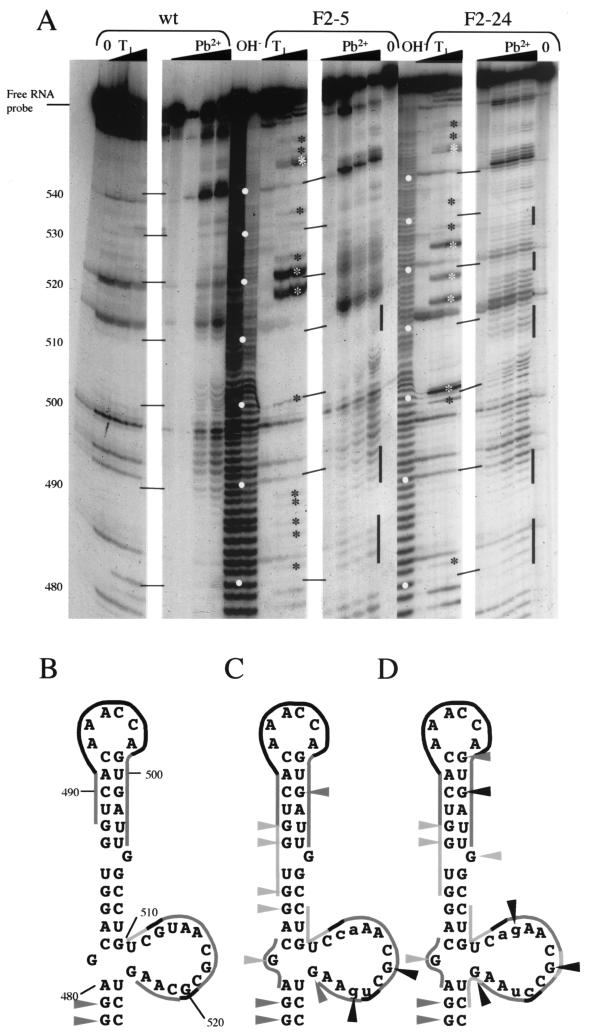
Translation of poliovirus RNA is driven by an internal ribosome entry segment (IRES) present in the 5' noncoding region of the genomic RNA. This IRES is structured into several domains, including domain V, which contains a large lateral bulge-loop whose predicted secondary structure is unclear. The primary sequence of this bulge-loop is strongly conserved within enteroviruses and rhinoviruses: it encompasses two GNAA motifs which could participate in intrabulge base pairing or (in one case) could be presented as a GNRA tetraloop. We have begun to address the question of the significance of the sequence conservation observed among enterovirus reference strains and field isolates by using a comprehensive site-directed mutagenesis program targeted to these two GNAA motifs. Mutants were analyzed functionally in terms of (i) viability and growth kinetics in both HeLa and neuronal cell lines, (ii) structural analyses by biochemical probing of the RNA, and (iii) translation initiation efficiencies in vitro in rabbit reticulocyte lysates supplemented with HeLa or neuronal cell extracts. Phenotypic analyses showed that only viruses with both GNAA motifs destroyed were significantly affected in their growth capacities, which correlated with in vitro translation defects. The phenotypic defects were strongly exacerbated in neuronal cells, where a temperature-sensitive phenotype could be revealed at between 37 and 39.5 degrees C. Biochemical probing of mutated domain V, compared to the wild type, demonstrated that such mutations lead to significant structural perturbations. Interestingly, revertant viruses possessed compensatory mutations which were distant from the primary mutations in terms of sequence and secondary structure, suggesting that intradomain tertiary interactions could exist within domain V of the IRES.
Figures
Similar articles
-
Attenuation stem-loop lesions in the 5' noncoding region of poliovirus RNA: neuronal cell-specific translation defects.J Virol. 1996 Mar;70(3):1467-74. doi: 10.1128/JVI.70.3.1467-1474.1996. J Virol. 1996. PMID: 8627664 Free PMC article.
-
Minimum internal ribosome entry site required for poliovirus infectivity.J Virol. 1993 Dec;67(12):7461-71. doi: 10.1128/JVI.67.12.7461-7471.1993. J Virol. 1993. PMID: 8230467 Free PMC article.
-
Role of GNRA motif mutations within stem-loop V of internal ribosome entry segment in coxsackievirus B3 molecular attenuation.J Mol Microbiol Biotechnol. 2008;14(4):147-56. doi: 10.1159/000107369. Epub 2007 Aug 13. J Mol Microbiol Biotechnol. 2008. PMID: 17693702
-
Effects of the Sabin-like mutations in domain V of the internal ribosome entry segment on translational efficiency of the Coxsackievirus B3.Mol Genet Genomics. 2006 Oct;276(4):402-12. doi: 10.1007/s00438-006-0155-3. Epub 2006 Aug 15. Mol Genet Genomics. 2006. PMID: 16909284
-
Pyrimidine-rich region mutations compensate for a stem-loop V lesion in the 5' noncoding region of poliovirus genomic RNA.Virology. 1999 Nov 25;264(2):385-97. doi: 10.1006/viro.1999.9981. Virology. 1999. PMID: 10562500
Cited by
-
Molecular Analysis of RNA-RNA Interactions between 5' and 3' Untranslated Regions during the Initiation of Translation of a Cardiovirulent and a Live-Attenuated Coxsackievirus B3 Strains.Int J Mol Sci. 2013 Feb 25;14(3):4525-44. doi: 10.3390/ijms14034525. Int J Mol Sci. 2013. PMID: 23439556 Free PMC article.
-
A mouse-adapted enterovirus 71 strain causes neurological disease in mice after oral infection.J Virol. 2004 Aug;78(15):7916-24. doi: 10.1128/JVI.78.15.7916-7924.2004. J Virol. 2004. PMID: 15254164 Free PMC article.
-
Comparative analysis of the large fragment of the 5' untranslated region (LF-5' UTR) of serotype A foot-and-mouth disease virus field isolates from India.Virus Genes. 2009 Aug;39(1):81-9. doi: 10.1007/s11262-009-0366-x. Epub 2009 May 15. Virus Genes. 2009. PMID: 19444600
-
Role of RNA structure motifs in IRES-dependent translation initiation of the coxsackievirus B3: new insights for developing live-attenuated strains for vaccines and gene therapy.Mol Biotechnol. 2013 Oct;55(2):179-202. doi: 10.1007/s12033-013-9674-4. Mol Biotechnol. 2013. PMID: 23881360 Review.
-
Correlation of mutations and recombination with growth kinetics of poliovirus vaccine strains.Eur J Clin Microbiol Infect Dis. 2010 Dec;29(12):1513-23. doi: 10.1007/s10096-010-1033-9. Epub 2010 Sep 5. Eur J Clin Microbiol Infect Dis. 2010. PMID: 20820837
References
-
- Andino, R., G. E. Rieckhof, and D. Baltimore. 1991. A functional ribonucleoprotein complex forms around the 5′end of poliovirus RNA. Cell 63:369-380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
