

Identification of conserved and variable structures in the human immunodeficiency virus gp120 glycoprotein of importance for CXCR4 binding
- PMID: 12368322
- PMCID: PMC136628
- DOI: 10.1128/jvi.76.21.10791-10800.2002
Identification of conserved and variable structures in the human immunodeficiency virus gp120 glycoprotein of importance for CXCR4 binding
Abstract
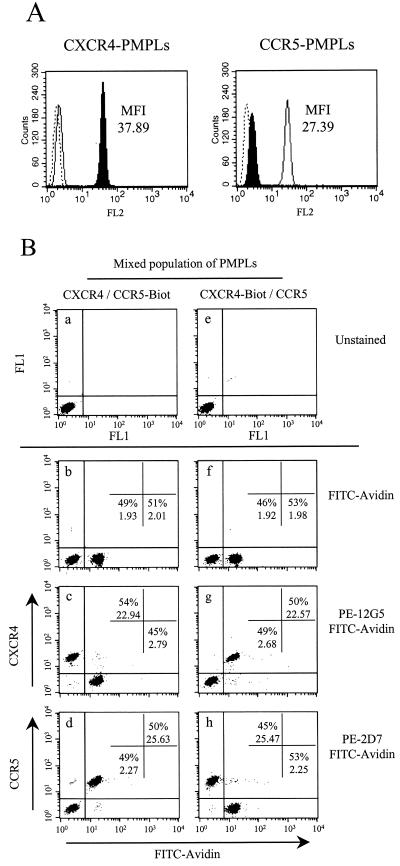
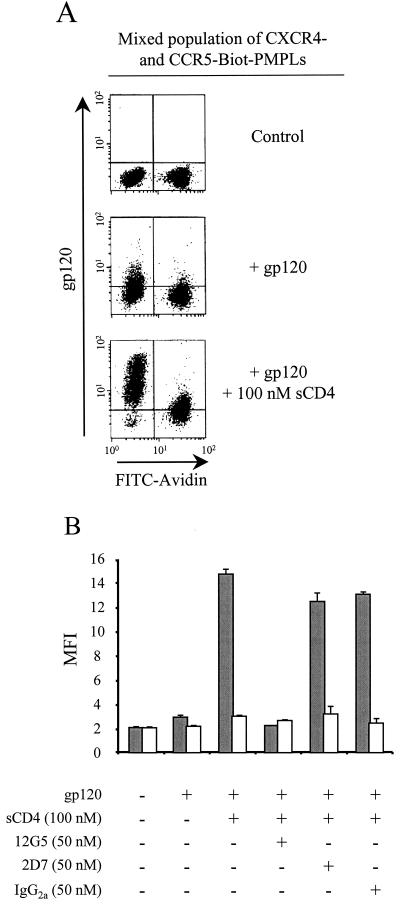
CD4 and the chemokine receptors, CXCR4 and CCR5, serve as receptors for human immunodeficiency virus type 1 (HIV-1). Binding of the HIV-1 gp120 envelope glycoprotein to the chemokine receptors normally requires prior interaction with CD4. Mapping the determinants on gp120 for the low-affinity interaction with CXCR4 has been difficult due to the nonspecific binding of this viral glycoprotein to cell surfaces. Here we examine the binding of a panel of gp120 mutants to paramagnetic proteoliposomes displaying CXCR4 on their surfaces. We show that the gp120 beta19 strand and third variable (V3) loop contain residues important for CXCR4 interaction. Basic residues from both elements, as well as a conserved hydrophobic residue at the V3 tip, contribute to CXCR4 binding. Removal of the gp120 V1/V2 variable loops allows the envelope glycoprotein to bind CXCR4 in a CD4-independent manner. These results indicate that although some variable gp120 residues contribute to the specific binding to CCR5 or CXCR4, gp120 elements common to CXCR4- or CCR5-using strains are involved in the interaction with both coreceptors.
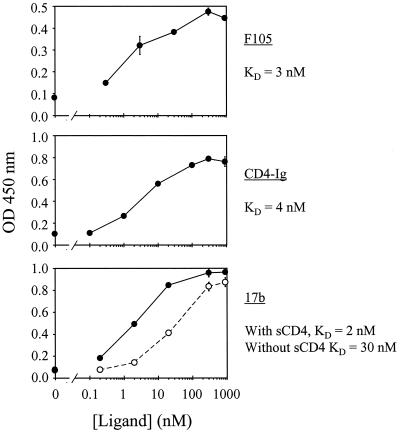
Figures
References
-
- Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger. 1996. CC CKR5: a RANTES, MIP-1α, MIP-1β receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272:1955-1958. - PubMed
-
- Babcock, G. J., T. Mirzabekov, W. Wojtowicz, and J. Sodroski. 2001. Ligand binding characteristics of CXCR4 incorporated into paramagnetic proteoliposomes. J. Biol. Chem. 276:38433-38440. - PubMed
-
- Blanpain, C., B. Lee, J. Vakili, B. J. Doranz, C. Govaerts, I. Migeotte, M. Sharron, V. Dupriez, G. Vassart, R. W. Doms, and M. Parmentier. 1999. Extracellular cysteines of CCR5 are required for chemokine binding, but dispensable for HIV-1 coreceptor activity. J. Biol. Chem. 274:18902-18908. - PubMed
-
- Brelot, A., N. Heveker, M. Montes, and M. Alizon. 2000. Identification of residues of CXCR4 critical for human immunodeficiency virus coreceptor and chemokine receptor activities. J. Biol. Chem. 275:23736-23744. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
