

Severe developmental defects, hypersensitivity to DNA-damaging agents, and lengthened telomeres in Arabidopsis MRE11 mutants
- PMID: 12368497
- PMCID: PMC151228
- DOI: 10.1105/tpc.005959
Severe developmental defects, hypersensitivity to DNA-damaging agents, and lengthened telomeres in Arabidopsis MRE11 mutants
Abstract
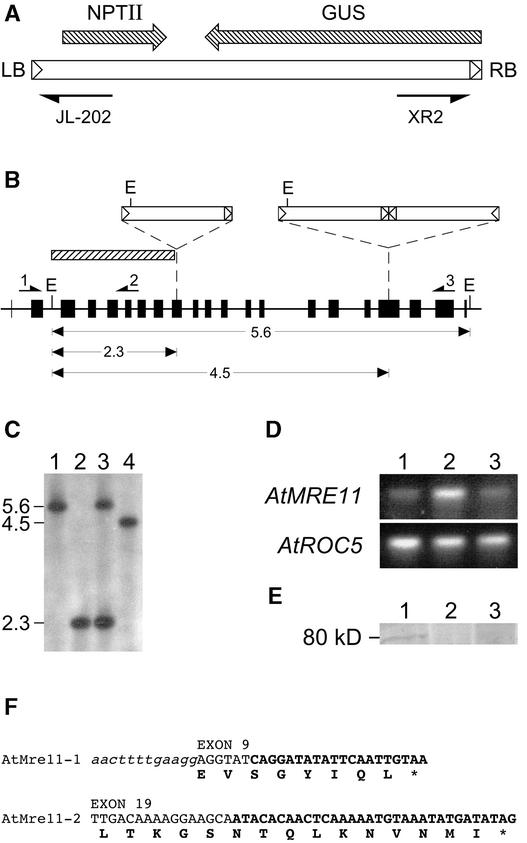
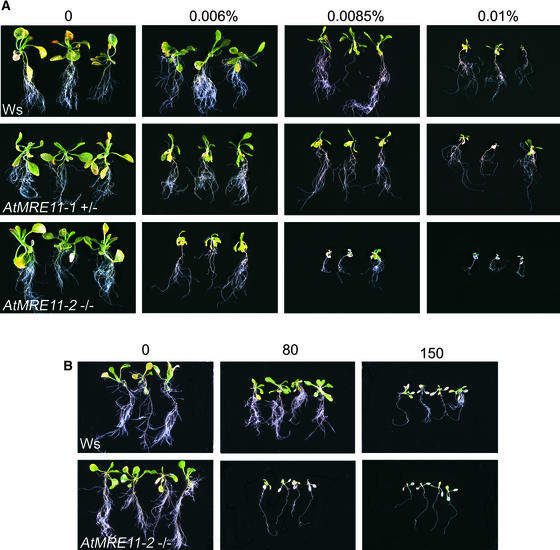
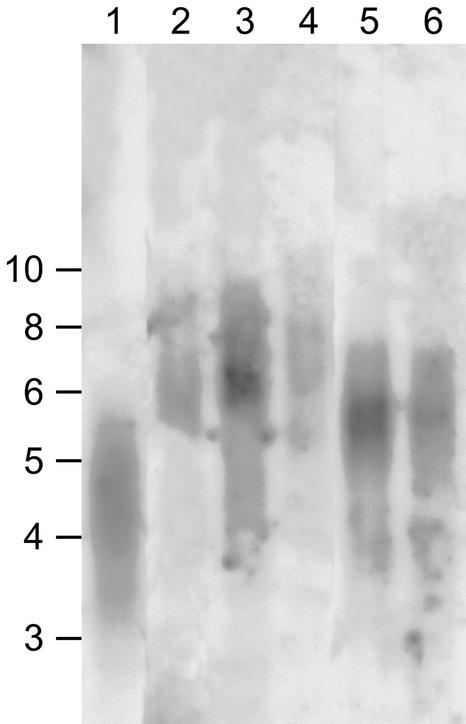
The Mre11 protein is essential for the long-term genetic stability of the cell and acts to ensure the efficient repair of DNA damage. Vertebrate cells lacking Mre11 function are not viable. However, we report here that this is not the case in the model plant Arabidopsis. We have isolated two different Arabidopsis lines containing a T-DNA copy integrated at a different point in the MRE11 gene (AtMRE11). Both mutant plant lines were hypersensitive to DNA-damaging treatments but exhibited strikingly different developmental phenotypes. Furthermore, we also observed lengthened telomeres in these plant lines, showing that AtMre11 is involved in telomere maintenance. Thus, the lines we have isolated are unique tools with which to study in detail the role of AtMre11 in the mature plant.
Figures


Similar articles
-
Arabidopsis thalianaMRE11 is essential for activation of cell cycle arrest, transcriptional regulation and DNA repair upon the induction of double-stranded DNA breaks.Plant Biol (Stuttg). 2016 Jul;18(4):681-94. doi: 10.1111/plb.12453. Epub 2016 Apr 13. Plant Biol (Stuttg). 2016. PMID: 27007017
-
Disruption of the Arabidopsis RAD50 gene leads to plant sterility and MMS sensitivity.Plant J. 2001 Jan;25(1):31-41. doi: 10.1046/j.1365-313x.2001.00928.x. Plant J. 2001. PMID: 11169180
-
Increased telomere length and hypersensitivity to DNA damaging agents in an Arabidopsis KU70 mutant.Nucleic Acids Res. 2002 Aug 1;30(15):3395-400. doi: 10.1093/nar/gkf445. Nucleic Acids Res. 2002. PMID: 12140324 Free PMC article.
-
Arabidopsis TAF1 is an MRE11-interacting protein required for resistance to genotoxic stress and viability of the male gametophyte.Plant J. 2015 Nov;84(3):545-57. doi: 10.1111/tpj.13020. Epub 2015 Oct 8. Plant J. 2015. PMID: 26358508 Free PMC article.
-
DNA repair mutants in higher eukaryotes.J Cell Sci Suppl. 1987;6:61-82. doi: 10.1242/jcs.1984.supplement_6.4. J Cell Sci Suppl. 1987. PMID: 3308925 Review.
Cited by
-
Telomere- and Telomerase-Associated Proteins and Their Functions in the Plant Cell.Front Plant Sci. 2016 Jun 28;7:851. doi: 10.3389/fpls.2016.00851. eCollection 2016. Front Plant Sci. 2016. PMID: 27446102 Free PMC article. Review.
-
Suppression of RICE TELOMERE BINDING PROTEIN 1 results in severe and gradual developmental defects accompanied by genome instability in rice.Plant Cell. 2007 Jun;19(6):1770-81. doi: 10.1105/tpc.107.051953. Epub 2007 Jun 22. Plant Cell. 2007. PMID: 17586654 Free PMC article.
-
Molecular analysis of telomere fusions in Arabidopsis: multiple pathways for chromosome end-joining.EMBO J. 2004 Jun 2;23(11):2304-13. doi: 10.1038/sj.emboj.7600236. Epub 2004 May 13. EMBO J. 2004. PMID: 15141167 Free PMC article.
-
A DNA2 Homolog Is Required for DNA Damage Repair, Cell Cycle Regulation, and Meristem Maintenance in Plants.Plant Physiol. 2016 May;171(1):318-33. doi: 10.1104/pp.16.00312. Epub 2016 Mar 7. Plant Physiol. 2016. PMID: 26951435 Free PMC article.
-
MRE11 is required for homologous synapsis and DSB processing in rice meiosis.Chromosoma. 2013 Oct;122(5):363-76. doi: 10.1007/s00412-013-0421-1. Epub 2013 Jun 22. Chromosoma. 2013. PMID: 23793712
References
-
- Carney, J.P., Maser, R.S., Olivares, H., Davies, E.M., Le Beau, M., Yates, J.R., Hays, L., Morgan, J.F., and Petrini, J.H. (1998). The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: Linkage of double-strand break repair to the cellular DNA damage response. Cell 93, 477–486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
