

CD4 effector T cell subsets in the response to influenza: heterogeneity, migration, and function
- PMID: 12370257
- PMCID: PMC2194021
- DOI: 10.1084/jem.20021052
CD4 effector T cell subsets in the response to influenza: heterogeneity, migration, and function
Abstract
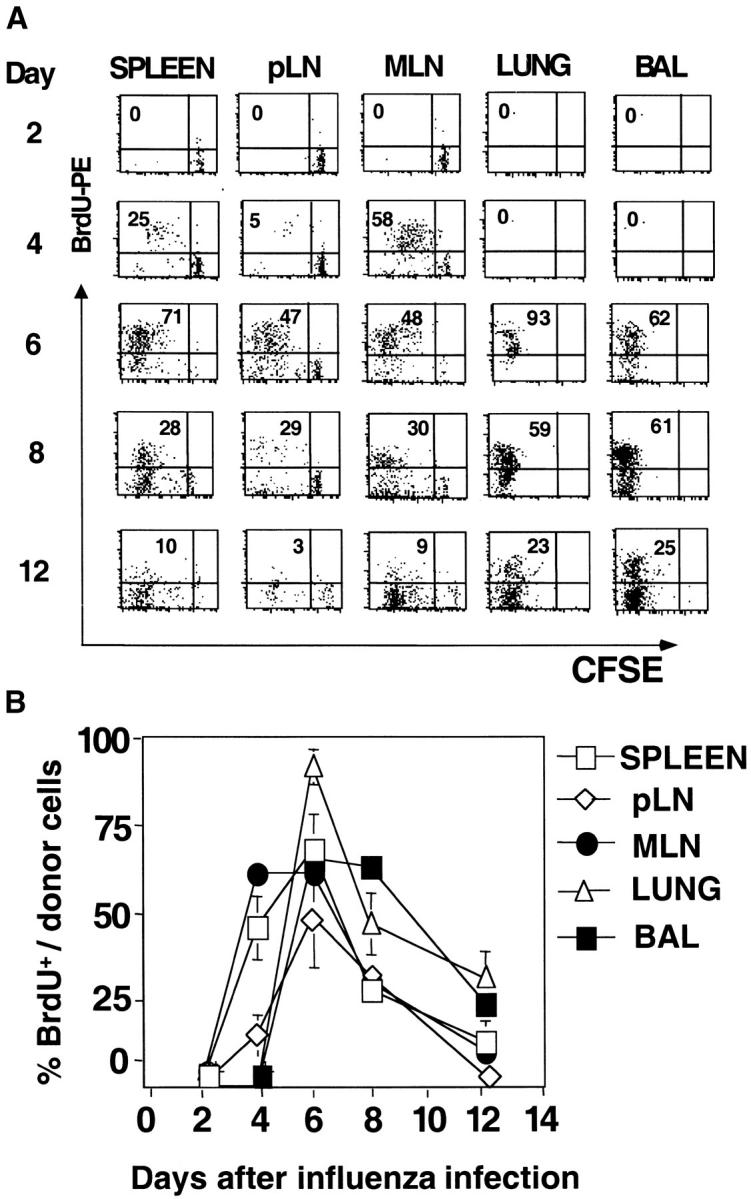
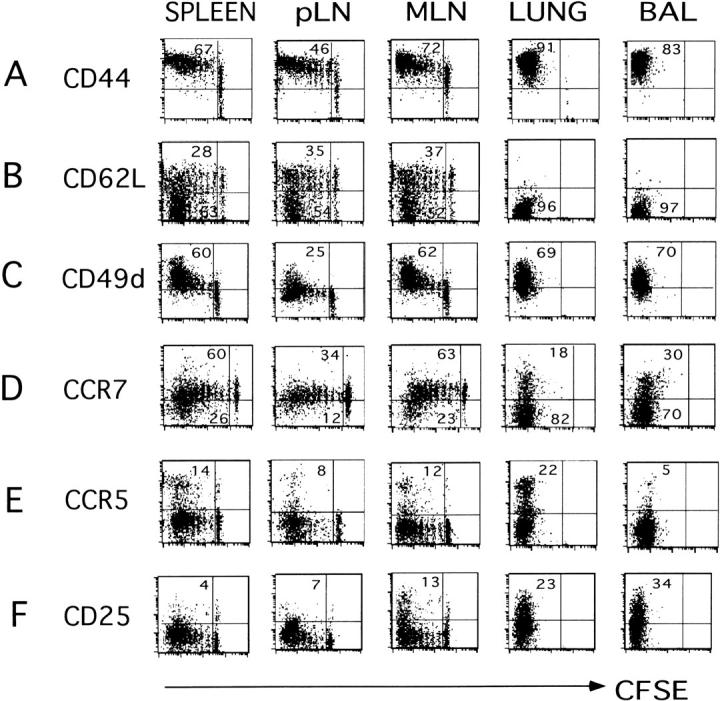
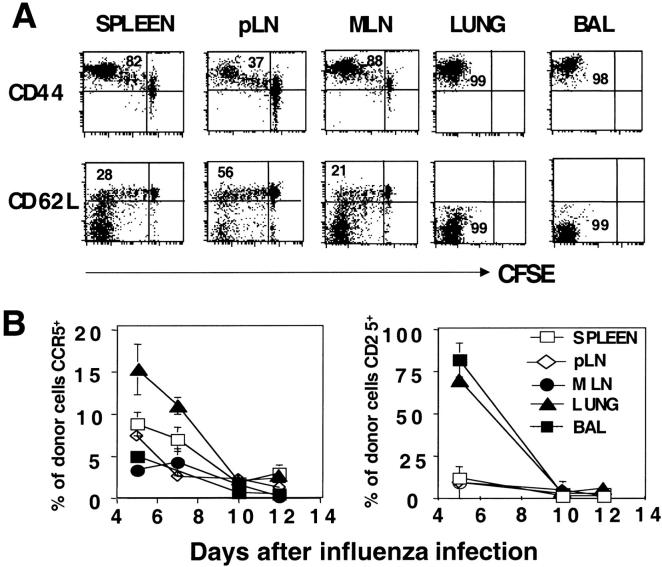
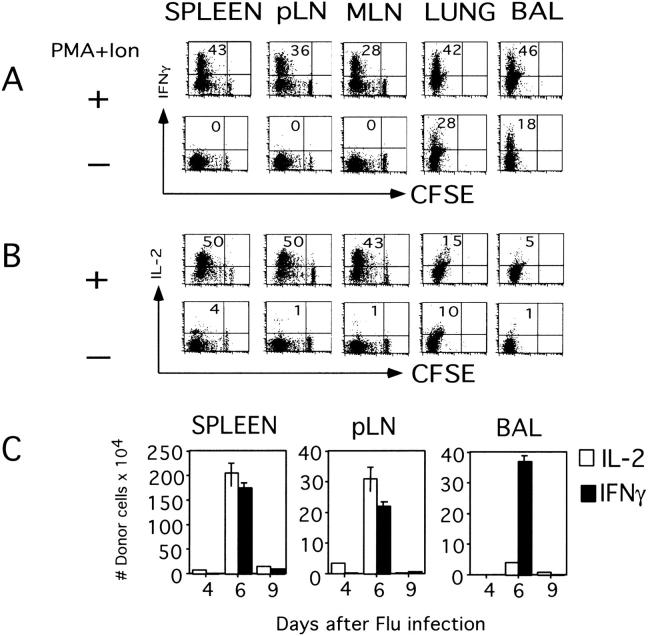
The immune response of naive CD4 T cells to influenza virus is initiated in the draining lymph nodes and spleen, and only after effectors are generated do antigen-specific cells migrate to the lung which is the site of infection. The effector cells generated in secondary organs appear as multiple subsets which are a heterogeneous continuum of cells in terms of number of cell divisions, phenotype and function. The effector cells that migrate to the lung constitute the more differentiated of the total responding population, characterized by many cell divisions, loss of CD62L, down-regulation of CCR7, stable expression of CD44 and CD49d, and transient expression of CCR5 and CD25. These cells also secrete high levels of interferon gamma and reduced levels of interleukin 2 relative to those in the secondary lymphoid organs. The response declines rapidly in parallel with viral clearance, but a spectrum of resting cell subsets reflecting the pattern at the peak of response is retained, suggesting that heterogeneous effector populations may give rise to corresponding memory populations. These results reveal a complex response, not an all-or-none one, which results in multiple effector phenotypes and implies that effector cells and the memory cells derived from them can display a broad spectrum of functional potentials.
Figures

References
-
- Jenkins, M.K., A. Khoruts, E. Ingulli, D.L. Mueller, J.S. McSorley, R.L. Reinhardt, A. Itano, and K.A. Pape. 2001. In vivo activation of antigen-specific CD4 T cells. Annu. Rev. Immunol. 19:23–45. - PubMed
-
- Mercado, R., S. Vijh, S.E. Allen, K. Kerksiek, I.M. Pilip, and E.G. Pamer. 2000. Early programming of T cell populations responding to bacterial infection. J. Immunol. 165:6833–6839. - PubMed
-
- Van Stipdonk, M.J.B., E.E. Lemmens, and S.P. Schoenberger. 2001. Naive CTLs require a single brief period of antigenic stimulation for clonal expansion and differentiation. Nat. Immunol. 2:423–429. - PubMed
-
- Laouar, Y., and I.N. Crispe. 2000. Functional flexibility in T cells: Independent regulation of CD4+ T cell proliferation and effector function in vivo. Immunity. 13:291–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
