

Ecto-5'-nucleotidase (CD73) regulation by hypoxia-inducible factor-1 mediates permeability changes in intestinal epithelia
- PMID: 12370277
- PMCID: PMC151145
- DOI: 10.1172/JCI15337
Ecto-5'-nucleotidase (CD73) regulation by hypoxia-inducible factor-1 mediates permeability changes in intestinal epithelia
Abstract
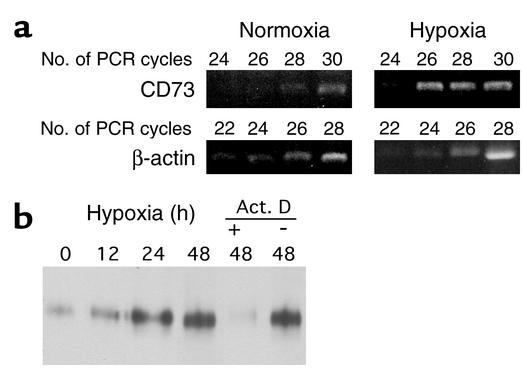
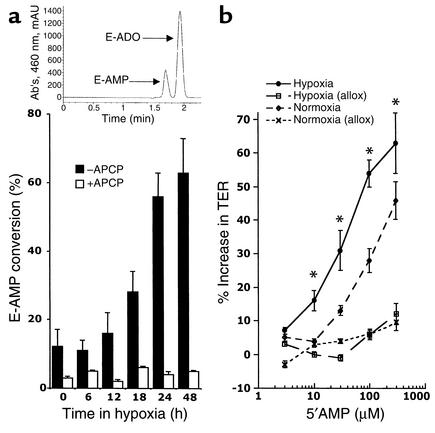
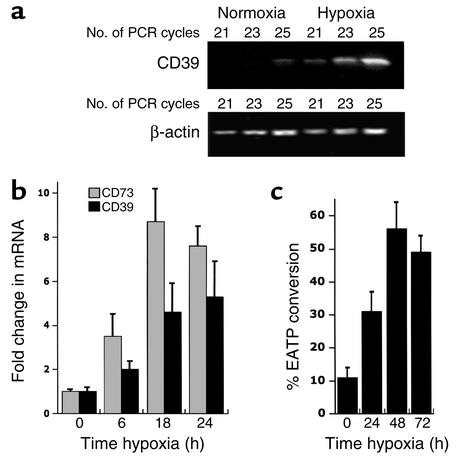
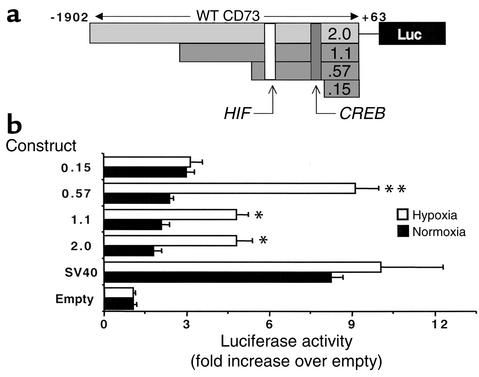
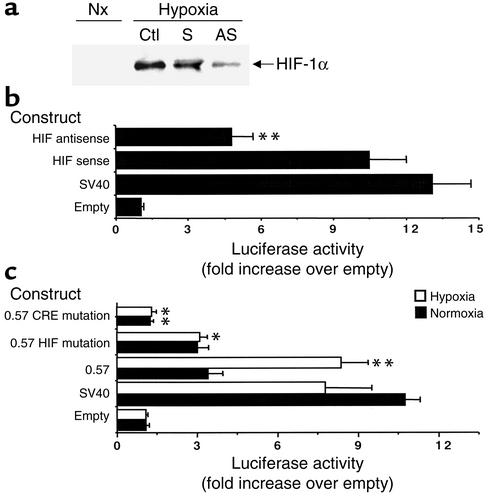
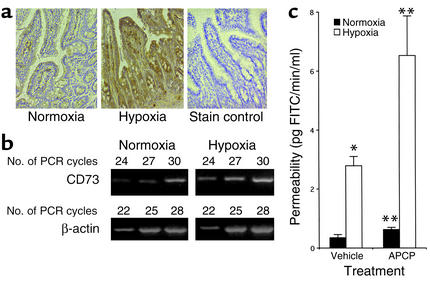
Under conditions of limited oxygen availability (hypoxia), multiple cell types release adenine nucleotides in the form of ATP, ADP, and AMP. Extracellular AMP is metabolized to adenosine by surface-expressed ecto-5'-nucleotidase (CD73) and subsequently activates surface adenosine receptors regulating endothelial and epithelial barrier function. Therefore, we hypothesized that hypoxia transcriptionally regulates CD73 expression. Microarray RNA analysis revealed an increase in CD73 and ecto-apyrase CD39 in hypoxic epithelial cells. Metabolic studies of CD39/CD73 function in intact epithelia revealed that hypoxia enhances CD39/CD73 function as much as 6 +/- 0.5-fold over normoxia. Examination of the CD73 gene promoter identified at least one binding site for hypoxia-inducible factor-1 (HIF-1) and inhibition of HIF-1alpha expression by antisense oligonucleotides resulted in significant inhibition of hypoxia-inducible CD73 expression. Studies using luciferase reporter constructs revealed a significant increase in activity in cells subjected to hypoxia, which was lost in truncated constructs lacking the HIF-1 site. Mutagenesis of the HIF-1alpha binding site resulted in a nearly complete loss of hypoxia-inducibility. In vivo studies in a murine hypoxia model revealed that hypoxia-induced CD73 may serve to protect the epithelial barrier, since the CD73 inhibitor alpha,beta-methylene ADP promotes increased intestinal permeability. These results identify an HIF-1-dependent regulatory pathway for CD73 and indicate the likelihood that CD39/CD73 protects the epithelial barrier during hypoxia.
Figures
References
-
- Pearson JD, Gordon JL. Nucleotide metabolism by endothelium. Annu Rev Physiol. 1985;47:617–627. - PubMed
-
- Zimmermann H, Braun N. Ecto-nucleotidases: molecular structures, catalytic properties, and functional roles in the nervous system. Prog Brain Res. 1999;120:371–385. - PubMed
-
- Stiles GL. Adenosine receptors. J Biol Chem. 1992;267:6451–6454. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
