

Presenilins mediate a dual intramembranous gamma-secretase cleavage of Notch-1
- PMID: 12374741
- PMCID: PMC129073
- DOI: 10.1093/emboj/cdf541
Presenilins mediate a dual intramembranous gamma-secretase cleavage of Notch-1
Abstract
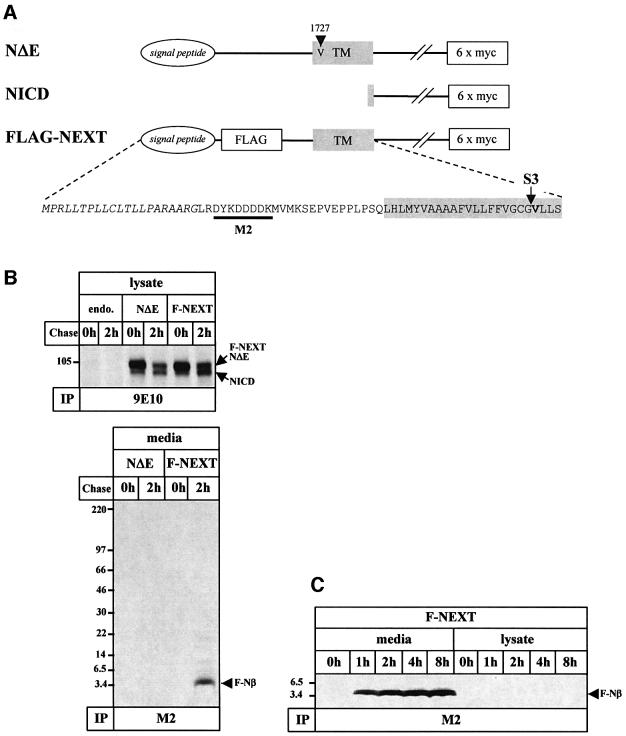
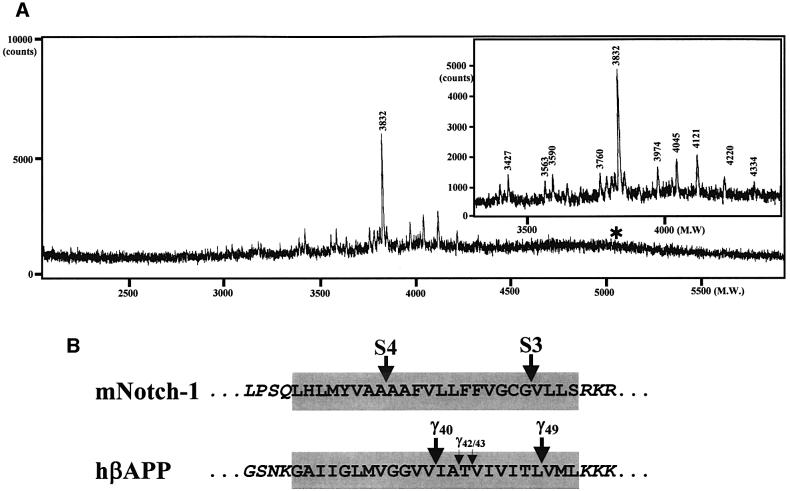
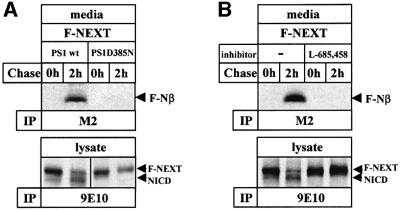
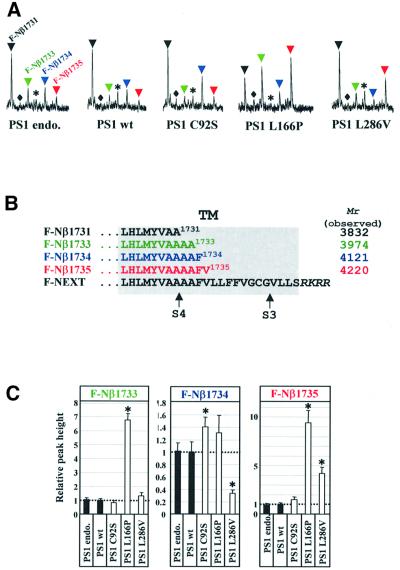
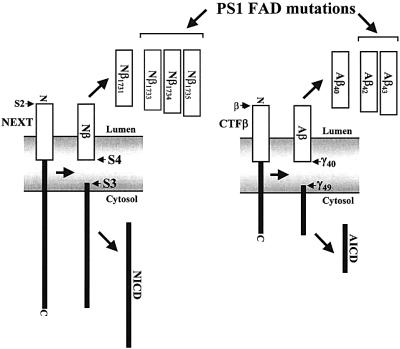
Following ectodomain shedding, Notch-1 undergoes presenilin (PS)-dependent constitutive intramembranous endoproteolysis at site-3. This cleavage is similar to the PS-dependent gamma-secretase cleavage of the beta-amyloid precursor protein (betaAPP). However, topological differences in cleavage resulting in amyloid beta-peptide (Abeta) or the Notch-1 intracellular domain (NICD) indicated independent mechanisms of proteolytic cleavage. We now demonstrate the secretion of an N-terminal Notch-1 Abeta-like fragment (Nbeta). Analysis of Nbeta by MALDI-TOF MS revealed that Nbeta is cleaved at a novel site (site-4, S4) near the middle of the transmembrane domain. Like the corresponding cleavage of betaAPP at position 40 and 42 of the Abeta domain, S4 cleavage is PS dependent. The precision of this cleavage is affected by familial Alzheimer's disease-associated PS1 mutations similar to the pathological endoproteolysis of betaAPP. Considering these similarities between intramembranous processing of Notch and betaAPP, we conclude that these proteins are cleaved by a common mechanism utilizing the same protease, i.e. PS/gamma-secretase.
Figures
References
-
- Beher D., Wrigley,J.D., Nadin,A., Evin,G., Masters,C.L., Harrison,T., Castro,J.L. and Shearman,M.S. (2001) Pharmacological knock-down of the presenilin 1 heterodimer by a novel γ-secretase inhibitor: implications for presenilin biology. J. Biol. Chem., 276, 45394–45402. - PubMed
-
- Brou C. et al. (2000) A novel proteolytic cleavage involved in Notch signaling: the role of the disintegrin-metalloprotease TACE. Mol. Cell, 5, 207–216. - PubMed
-
- Cao X. and Sudhof,T.C. (2001) A transcriptionally active complex of APP with Fe65 and histone acetyltransferase Tip60. Science, 293, 115–120. - PubMed
-
- Capell A., Steiner,H., Romig,H., Keck,S., Baader,M., Grim,M.G., Baumeister,R. and Haass,C. (2000) Presenilin-1 differentially facilitates endoproteolysis of the β-amyloid precursor protein and Notch. Nat. Cell Biol., 2, 205–211. - PubMed
-
- Chung H.M. and Struhl,G. (2001) Nicastrin is required for presenilin-mediated transmembrane cleavage in Drosophila. Nat. Cell Biol., 3, 1129–1132. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
