

Inducible control of virulence gene expression in Listeria monocytogenes: temporal requirement of listeriolysin O during intracellular infection
- PMID: 12374827
- PMCID: PMC135386
- DOI: 10.1128/JB.184.21.5935-5945.2002
Inducible control of virulence gene expression in Listeria monocytogenes: temporal requirement of listeriolysin O during intracellular infection
Abstract
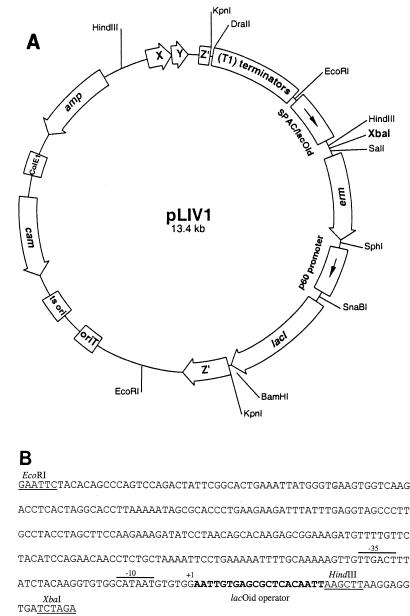
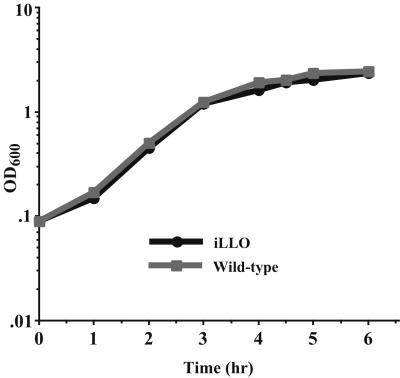
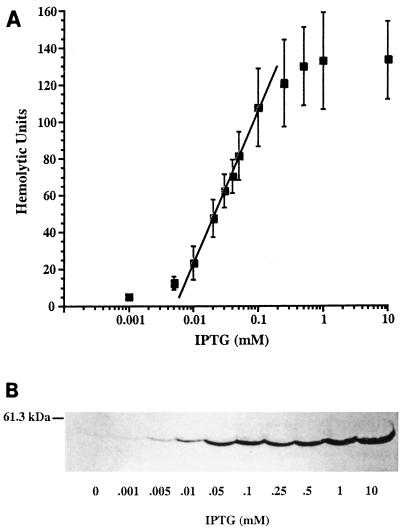
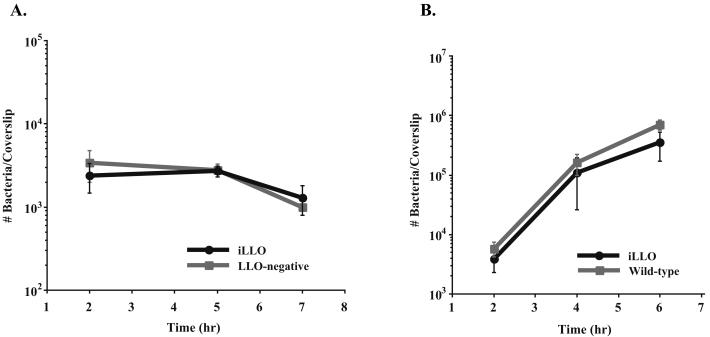
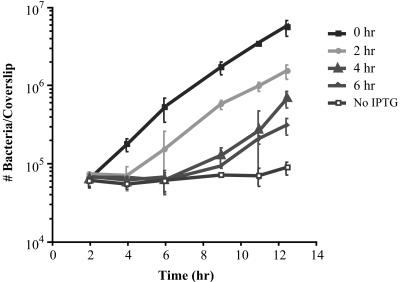
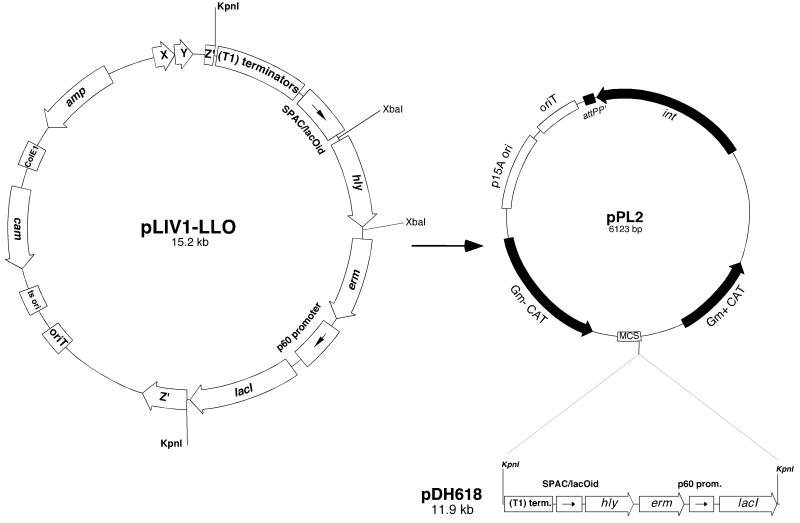
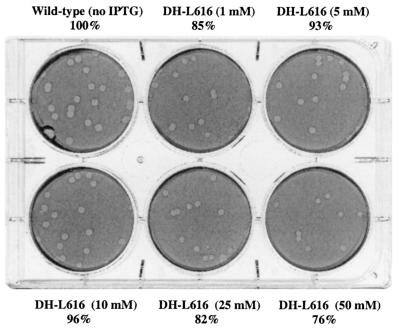
We have constructed a lac repressor/operator-based system to tightly regulate expression of bacterial genes during intracellular infection by Listeria monocytogenes. An L. monocytogenes strain was constructed in which expression of listeriolysin O was placed under the inducible control of an isopropyl-beta-D-thiogalactopyranoside (IPTG)-dependent promoter. Listeriolysin O (LLO) is a pore-forming cytolysin that mediates lysis of L. monocytogenes-containing phagosomes. Using hemolytic-activity assays and Western blot analysis, we demonstrated dose-dependent IPTG induction of LLO during growth in broth culture. Moreover, intracellular growth of the inducible-LLO (iLLO) strain in the macrophage-like cell line J774 was strictly dependent upon IPTG. We have further shown that iLLO bacteria trapped within primary phagocytic vacuoles can be induced to escape into the cytosol following addition of IPTG to the cell culture medium, thus yielding the ability to control bacterial escape from the phagosome and the initiation of intracellular growth. Using the iLLO strain in plaque-forming assays, we demonstrated an additional requirement for LLO in facilitating cell-to-cell spread in L2 fibroblasts, a nonprofessional phagocytic cell line. Furthermore, the efficiency of cell-to-cell spread of iLLO bacteria in L2 cells was IPTG dose dependent. The potential use of this system for determining the temporal requirements of additional virulence determinants of intracellular pathogenesis is discussed.
Figures

References
-
- Aureli, P., G. C. Fiorucci, D. Caroli, G. Marchiaro, O. Novara, L. Leone, and S. Salmaso. 2000. An outbreak of febrile gastroenteritis associated with corn contaminated by Listeria monocytogenes. N. Engl. J. Med. 342:1236-1241. - PubMed
-
- Bayley, H. 1997. Toxin structure: part of a hole? Curr. Biol. 7:R763-R767. - PubMed
-
- Beckwith, J. R., and D. Zisper. 1970. The lactose operon. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
-
- Biard, D. S. F., M. R. James, A. Cordier, and A. Sarasin. 1992. Regulation of the Escherichia coli lac operon expressed in human cells. Biochim. Biophys. 1130:68-74. - PubMed
-
- Bielecki, J., P. Youngman, P. Connelly, and D. A. Portnoy. 1990. Bacillus subtilis expressing a haemolysin gene from Listeria monocytogenes can grow in mammalian cells. Nature 345:175-176. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
