

The first archaeal ATP-dependent glucokinase, from the hyperthermophilic crenarchaeon Aeropyrum pernix, represents a monomeric, extremely thermophilic ROK glucokinase with broad hexose specificity
- PMID: 12374829
- PMCID: PMC135380
- DOI: 10.1128/JB.184.21.5955-5965.2002
The first archaeal ATP-dependent glucokinase, from the hyperthermophilic crenarchaeon Aeropyrum pernix, represents a monomeric, extremely thermophilic ROK glucokinase with broad hexose specificity
Abstract
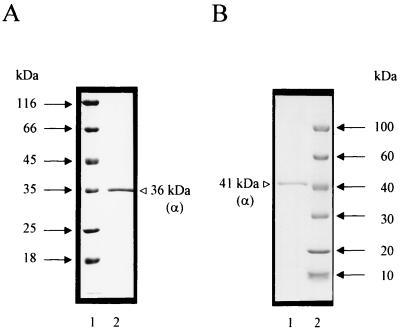
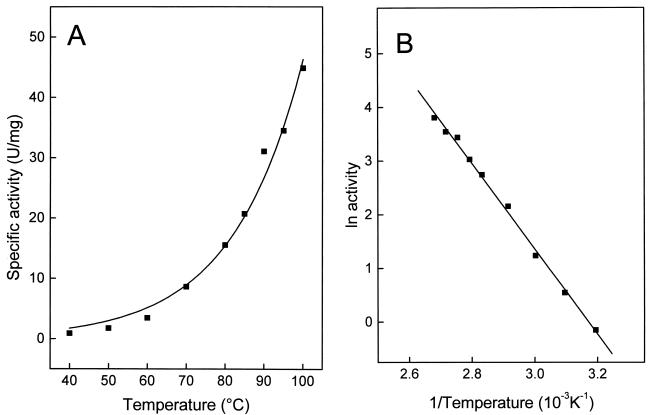
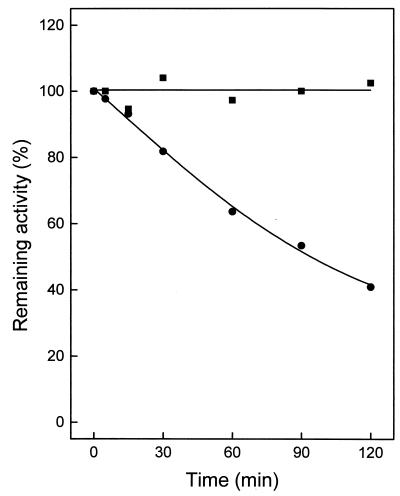
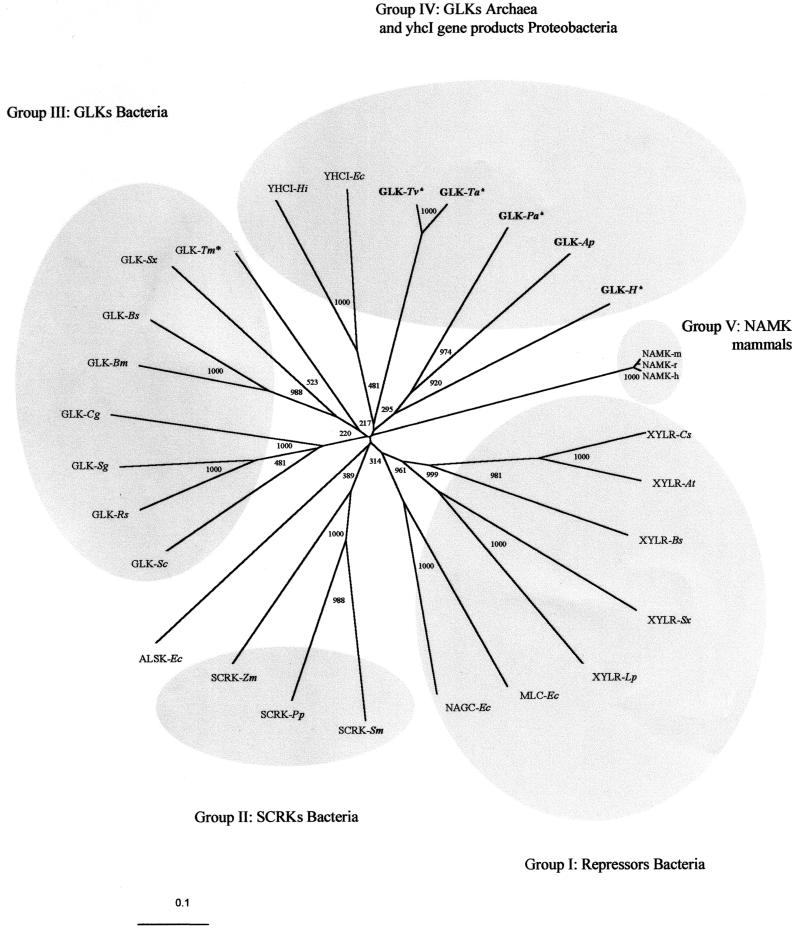
An ATP-dependent glucokinase of the hyperthermophilic aerobic crenarchaeon Aeropyrum pernix was purified 230-fold to homogeneity. The enzyme is a monomeric protein with an apparent molecular mass of about 36 kDa. The apparent K(m) values for ATP and glucose (at 90 degrees C and pH 6.2) were 0.42 and 0.044 mM, respectively; the apparent V(max) was about 35 U/mg. The enzyme was specific for ATP as a phosphoryl donor, but showed a broad spectrum for phosphoryl acceptors: in addition to glucose, which showed the highest catalytic efficiency (k(cat)/K(m)), the enzyme also phosphorylates glucosamin, fructose, mannose, and 2-deoxyglucose. Divalent cations were required for maximal activity: Mg(2+), which was most effective, could partially be replaced with Co(2+), Mn(2+), and Ni(2+). The enzyme had a temperature optimum of at least 100 degrees C and showed significant thermostability up to 100 degrees C. The coding function of open reading frame (ORF) APE2091 (Y. Kawarabayasi, Y. Hino, H. Horikawa, S. Yamazaki, Y. Haikawa, K. Jin-no, M. Takahashi, M. Sekine, S. Baba, A. Ankai, H. Kosugi, A. Hosoyama, S. Fukui, Y. Nagai, K. Nishijima, H. Nakazawa, M. Takamiya, S. Masuda, T. Funahashi, T. Tanaka, Y. Kudoh, J. Yamazaki, N. Kushida, A. Oguchi, and H. Kikuchi, DNA Res. 6:83-101, 145-152, 1999), previously annotated as gene glk, coding for ATP-glucokinase of A. pernix, was proved by functional expression in Escherichia coli. The purified recombinant ATP-dependent glucokinase showed a 5-kDa higher molecular mass on sodium dodecyl sulfate-polyacrylamide gel electrophoresis, but almost identical kinetic and thermostability properties in comparison to the native enzyme purified from A. pernix. N-terminal amino acid sequence of the native enzyme revealed that the translation start codon is a GTG 171 bp downstream of the annotated start codon of ORF APE2091. The amino acid sequence deduced from the truncated ORF APE2091 revealed sequence similarity to members of the ROK family, which comprise bacterial sugar kinases and transcriptional repressors. This is the first report of the characterization of an ATP-dependent glucokinase from the domain of Archaea, which differs from its bacterial counterparts by its monomeric structure and its broad specificity for hexoses.
Figures

References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403-410. - PubMed
-
- Arora, K. K., P. Shenbagamurthi, M. Fanciulli, and P. L. Pedersen. 1990. Glucose phosphorylation. Interaction of a 50-amino acid peptide of yeast hexokinase with trinitrophenyl ATP. J. Biol. Chem. 265:5324-5328. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
