

Down-regulation of TM29, a tomato SEPALLATA homolog, causes parthenocarpic fruit development and floral reversion
- PMID: 12376628
- PMCID: PMC166590
- DOI: 10.1104/pp.005223
Down-regulation of TM29, a tomato SEPALLATA homolog, causes parthenocarpic fruit development and floral reversion
Abstract
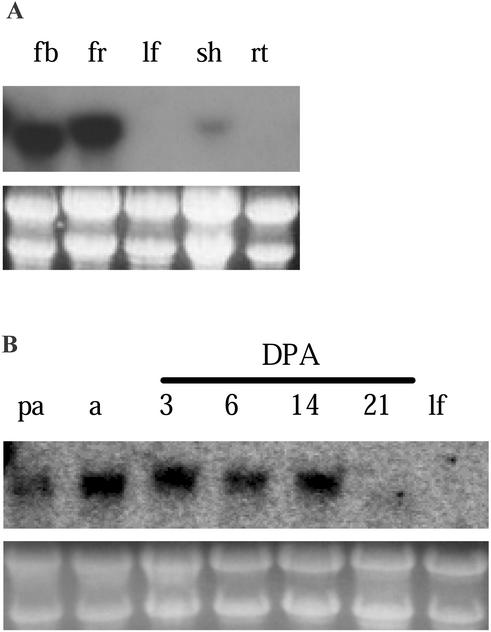
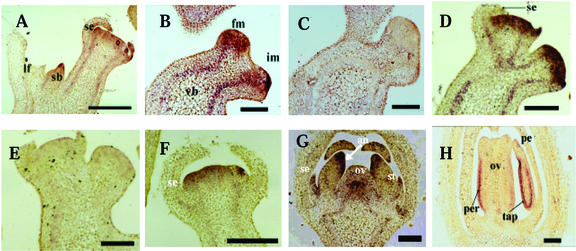
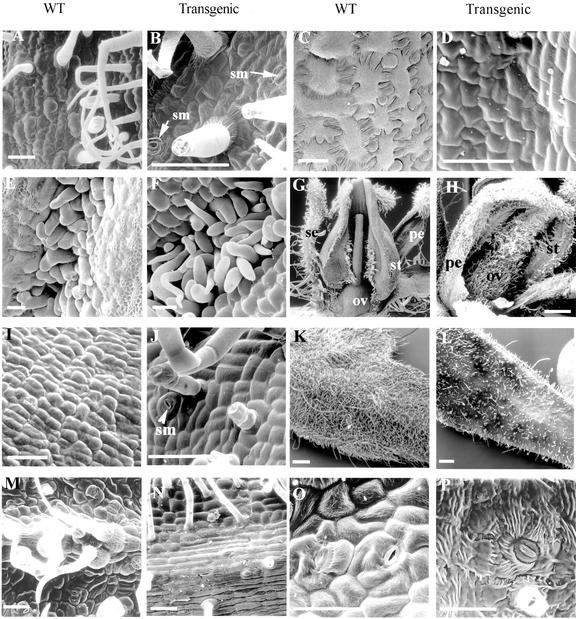
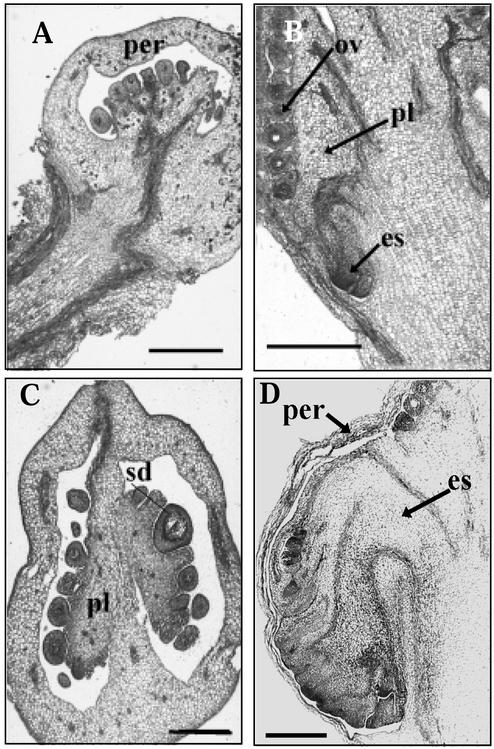
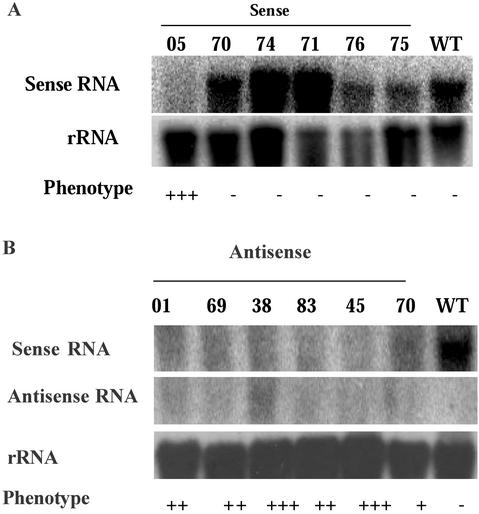
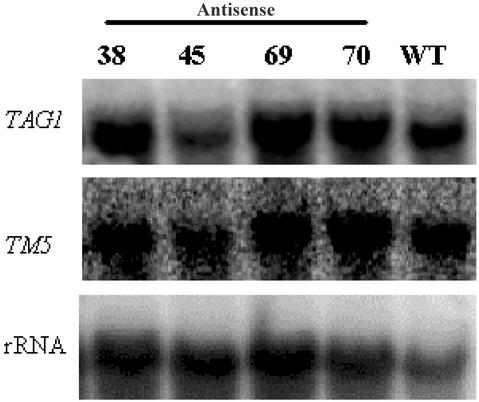
We have characterized the tomato (Lycopersicon esculentum Mill.) MADS box gene TM29 that shared a high amino acid sequence homology to the Arabidopsis SEP1, 2, and 3 (SEPALLATA1, 2, and 3) genes. TM29 showed similar expression profiles to SEP1, with accumulation of mRNA in the primordia of all four whorls of floral organs. In addition, TM29 mRNA was detected in inflorescence and vegetative meristems. To understand TM29 function, we produced transgenic tomato plants in which TM29 expression was down-regulated by either cosuppression or antisense techniques. These transgenic plants produced aberrant flowers with morphogenetic alterations in the organs of the inner three whorls. Petals and stamens were green rather than yellow, suggesting a partial conversion to a sepalloid identity. Stamens and ovaries were infertile, with the later developing into parthenocarpic fruit. Ectopic shoots with partially developed leaves and secondary flowers emerged from the fruit. These shoots resembled the primary transgenic flowers and continued to produce parthenocarpic fruit and additional ectopic shoots. Based on the temporal and spatial expression pattern and transgenic phenotypes, we propose that TM29 functions in floral organ development, fruit development, and maintenance of floral meristem identity in tomato.
Figures


References
-
- Allen KD, Sussex IM. Falsiflora and anantha control early stages of floral meristem development in tomato (Lycopersicon esculentum Mill.) Planta. 1996;200:254–264.
-
- Angenent GC, Franken J, Busscher M, Weiss D, van Tunen AJ. Co-suppression of the petunia homeotic gene fbp2 affects the identity of the generative meristem. Plant J. 1994;5:33–44. - PubMed
-
- Bowman JL, Smyth DR, Meyerowitz EM. Genetic interactions among floral homeotic genes of Arabidopsis. Development. 1991;112:1–20. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
