

Cytosolic free Ca(2+) changes and calpain activation are required for beta integrin-accelerated phagocytosis by human neutrophils
- PMID: 12379807
- PMCID: PMC2173489
- DOI: 10.1083/jcb.200206089
Cytosolic free Ca(2+) changes and calpain activation are required for beta integrin-accelerated phagocytosis by human neutrophils
Abstract
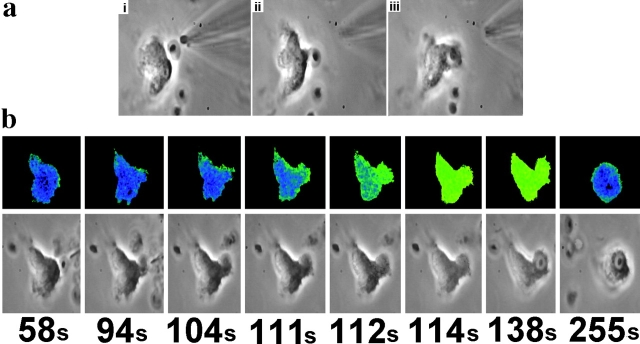
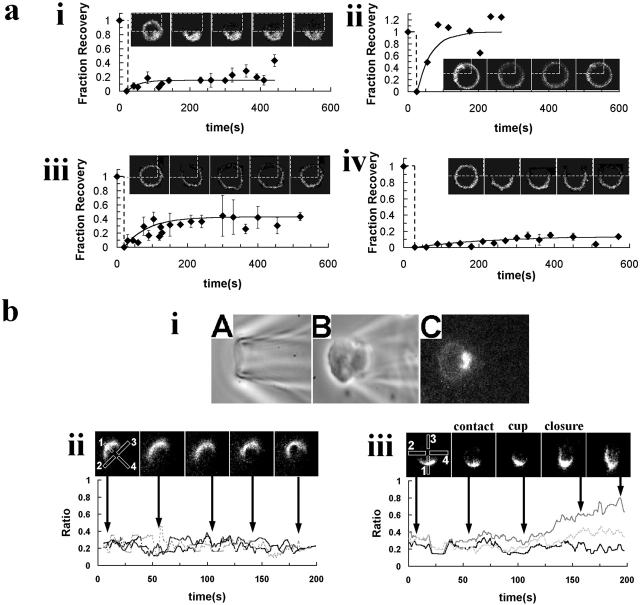
Phagocytosis of microbes coated with opsonins such as the complement component C3bi is the key activity of neutrophils. However, the mechanism by which opsonins enhance the rate of phagocytosis by these cells is unknown and has been difficult to study, partly because of the problem of observing and quantifying the events associated with phagocytosis. In this study, C3bi-opsonized particles were presented to neutrophils with a micromanipulator, so that the events of binding, pseudopod cup formation, engulfment, and completion of phagocytosis were clearly defined and distinguished from those involved with chemotaxis. Using this approach in combination with simultaneous phase contrast and Ca(2+) imaging, the temporal relationship between changes in cytosolic free Ca(2+) concentration and phagocytosis were correlated. Here we show that whereas small, localized Ca(2+) changes occur at the site of particle attachment and cup formation as a result of store release, rapid engulfment of the particle required a global change in cytosolic free Ca(2+) which resulted from Ca(2+) influx. This latter rise in cytosolic free Ca(2+) concentration also liberated a fraction of beta2 integrin receptors which were initially immobile on the neutrophil surface, as demonstrable by both fluorescence recovery after laser bleaching and by visualization of localized beta2 integrin labelling. Inhibitors of calpain activation prevented both the Ca(2+)-induced liberation of beta2 integrin and the rapid stage of phagocytosis, despite the persistence of the global Ca(2+) signal. Therefore, we propose that Ca(2+) activation of calpain causes beta2 integrin liberation, and that this signal plays a key role in the acceleration of beta2 integrin-mediated phagocytosis.
Figures








References
-
- Davies, E.V., and M.B. Hallett. 1999. High micromolar Ca2+ beneath the plasma membrane in stimulated neutrophils. Biochem. Biophys. Res. Commun. 248:679–683. - PubMed
-
- Hallett, M.B., E.V. Davies, and A.K. Campbell. 1990. Oxidase activation in individual neutrophils is dependent on the onset and magnitude of the Ca2+ signal. Cell Calcium. 11:655–663. - PubMed
-
- Hogg, N., and B. Leitinger. 2001. Shape and shift changes related to the function of leukocyte integrins LFA-1 and Mac-1. J. Leukoc. Biol. 69:893–898. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
