

IRES-driven translation is stimulated separately by the FMDV 3'-NCR and poly(A) sequences
- PMID: 12384586
- PMCID: PMC137133
- DOI: 10.1093/nar/gkf569
IRES-driven translation is stimulated separately by the FMDV 3'-NCR and poly(A) sequences
Abstract
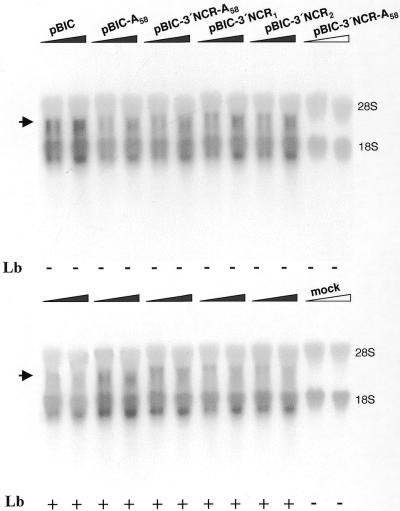
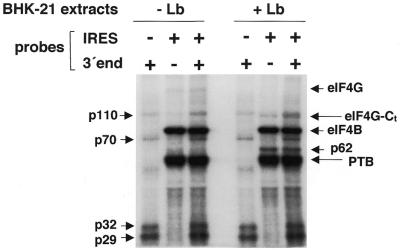
The 3' end region of foot-and-mouth disease virus (FMDV) consists of two distinct elements, a 90 nt untranslated region (3'-NCR) and a poly(A) tract. Removal of either the poly(A) tract or both the 3'-NCR and the poly(A) tract abrogated infectivity in susceptible cells in the context of a full-length cDNA clone. We have addressed the question of whether the impairment of RNA infectivity is related to defects at the translation level using a double approach. First, compared to the full-length viral RNA, removal of the 3' sequences reduced the efficiency of translation in vitro. Secondly, a stimulatory effect of the 3' end sequences on IRES-dependent translation was found in vivo using bicistronic constructs. RNAs carrying the FMDV 3' end sequences linked to the second cistron showed a significant stimulation of IRES-dependent translation, whereas cap-dependent translation was not affected. Remarkably, IRES-dependent stimulation exerted by the poly(A) tract or the 3'-NCR seems to be the result of two separate events, as the 3'-NCR alone enhanced IRES activity on its own. Under conditions of FMDV Lb protease-induced translation shut-off, the stimulation of IRES activity reached values above 6-fold in living cells. A northern blot analysis indicated that IRES stimulation was not the consequence of a change in the stability of the bicistronic RNA produced in transfected cells. Analysis of the RNA-binding proteins interacting with a mixture of 3' end and IRES probes showed an additive pattern. Altogether, our results strongly suggest that individual signals in the viral 3' end ensure stimulation of FMDV translation.
Figures
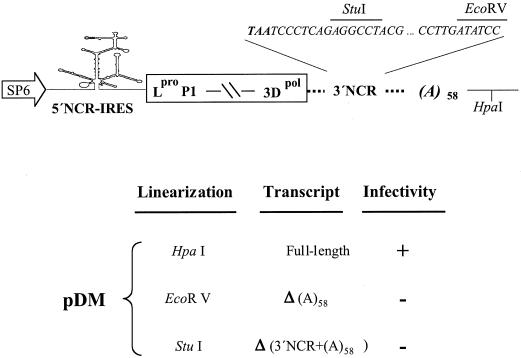
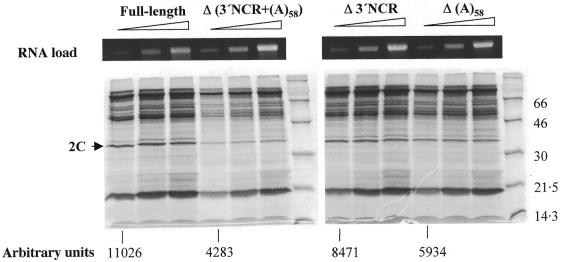
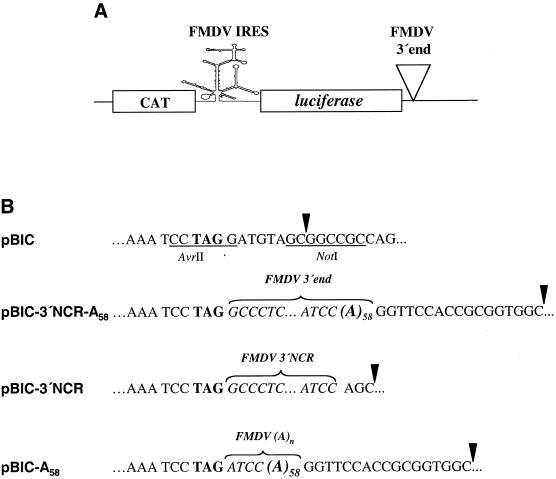
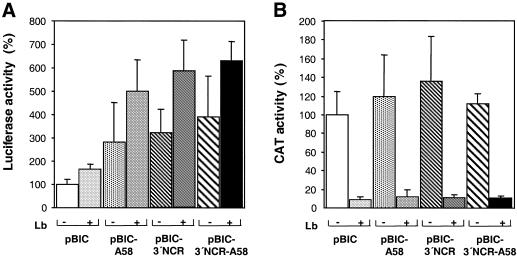
References
-
- Belsham G.J. and Jackson,R.J. (2000) Translation initiation on picornavirus RNA. In Sonenberg,N., Hershey,J.W.B. and Mathews,M.B. (eds), Translational Control of Gene Expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 869–900.
-
- Martínez-Salas E., Ramos,R., Lafuente,E. and López de Quinto,S. (2001) Functional interactions in internal translation initiation directed by viral and cellular IRES. J. Gen. Virol., 82, 973–984. - PubMed
-
- Hellen C.U. and Sarnow,P. (2001) Internal ribosome entry sites in eukaryotic mRNA molecules. Genes Dev., 15, 1593–1612. - PubMed
-
- Michel Y.M., Poncet,D., Piron,M., Kean,K.M. and Borman,A.M. (2000) Cap-poly(A) synergy in mammalian cell-free extracts. Investigation of the requirements for poly(A)-mediated stimulation of translation initiation. J. Biol. Chem., 275, 32268–32276. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
