

Induction of sporulation in Saccharomyces cerevisiae leads to the formation of N6-methyladenosine in mRNA: a potential mechanism for the activity of the IME4 gene
- PMID: 12384598
- PMCID: PMC137137
- DOI: 10.1093/nar/gkf573
Induction of sporulation in Saccharomyces cerevisiae leads to the formation of N6-methyladenosine in mRNA: a potential mechanism for the activity of the IME4 gene
Abstract
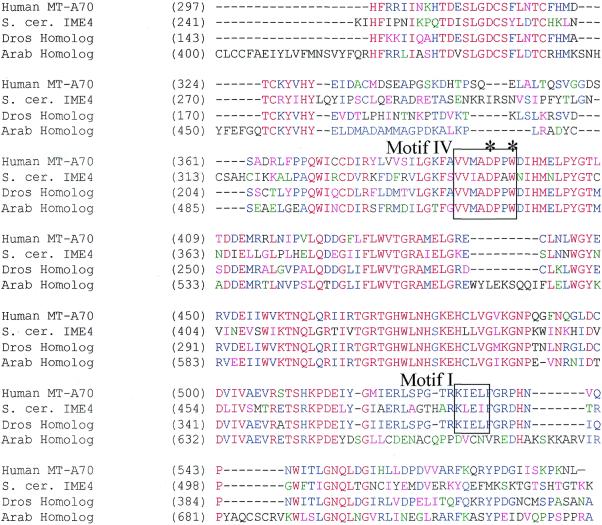
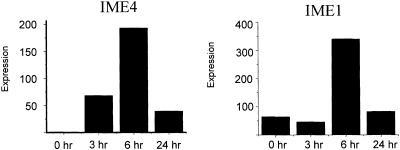
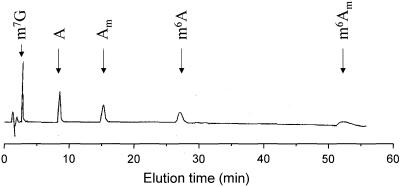
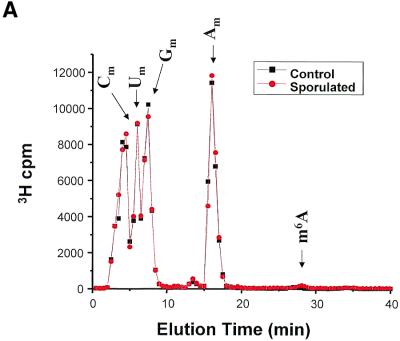
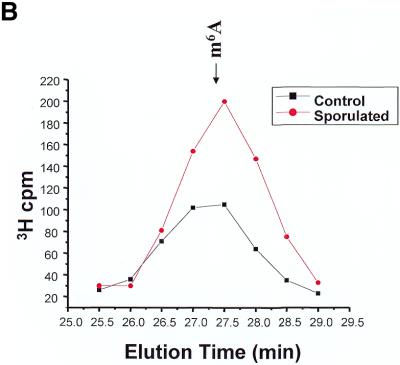
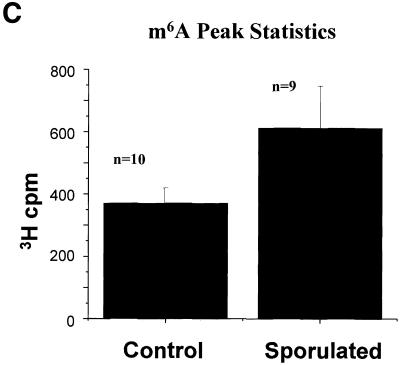
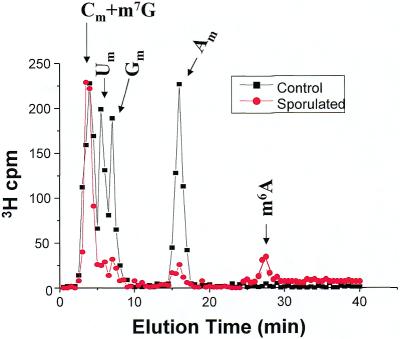
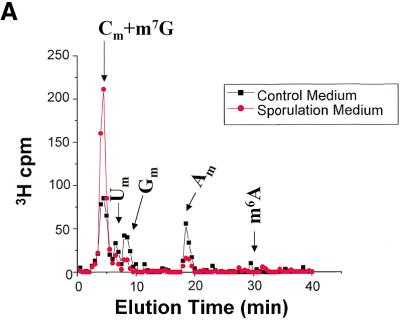
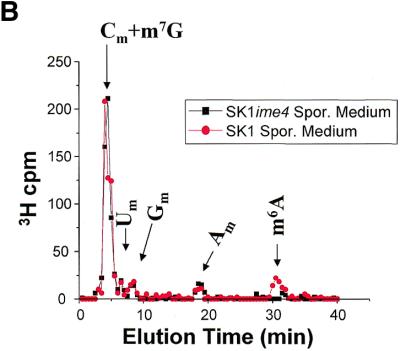
N6-methyladenosine (m6A) is present at internal sites in mRNA isolated from all higher eukaryotes, but has not previously been detected in the mRNA of the yeast Saccharomyces cerevisiae. This nucleoside modification occurs only in a sequence- specific context that appears to be conserved across diverse species. The function of this modification is not fully established, but there is some indirect evidence that m6A may play a role in the efficiency of mRNA splicing, transport or translation. The S.cerevisiae gene IME4, which is important for induction of sporulation, is very similar to the human gene MT-A70, which has been shown to be a critical subunit of the human mRNA [N6-adenosine]-methyltransferase. This observation led to the hypothesis that yeast sporulation may be dependent upon methylation of yeast mRNA, mediated by Ime4p. In this study we show that induction of sporulation leads to the appearance of low levels of m6A in yeast mRNA and that this modification requires IME4. Moreover, single amino acid substitutions in the putative catalytic residues of Ime4p lead to severe sporulation defects in a strain whose sporulation ability is completely dependent on this protein. Collectively, these data suggest very strongly that the activation of sporulation by Ime4p is the result of its proposed methyltransferase activity and provide the most direct evidence to date of a physiologic role of m6A in a gene regulatory pathway.
Figures
References
-
- Rottman F.M., Shatkin,A.J. and Perry,R.P. (1974) Sequences containing methylated nucleotides at the 5′-termini of messenger RNAs: possible implications for processing. Cell, 3, 197–199. - PubMed
-
- Reddy R., Singh,R. and Shimba,S. (1992) Methylated CAP structures in eukaryotic RNAs: structure, synthesis and function. Pharmacol. Ther., 54, 249–267. - PubMed
-
- Desrosiers R.C., Friderici,K.H. and Rottman,F.M. (1975) Characterization of Novikoff hepatoma mRNA methylation and heterogeneity in the methylated 5′ terminus. Biochemistry, 14, 4367–4374. - PubMed
-
- Perry R.P., Kelley,D.E., Friderici,K. and Rottman,F. (1975) The methylated constituents of L cell messenger RNA: evidence for an unusual cluster at the 5′ terminus. Cell, 4, 387–394. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
