

Analyses of single-copy Arabidopsis T-DNA-transformed lines show that the presence of vector backbone sequences, short inverted repeats and DNA methylation is not sufficient or necessary for the induction of transgene silencing
- PMID: 12384603
- PMCID: PMC137132
- DOI: 10.1093/nar/gkf568
Analyses of single-copy Arabidopsis T-DNA-transformed lines show that the presence of vector backbone sequences, short inverted repeats and DNA methylation is not sufficient or necessary for the induction of transgene silencing
Abstract
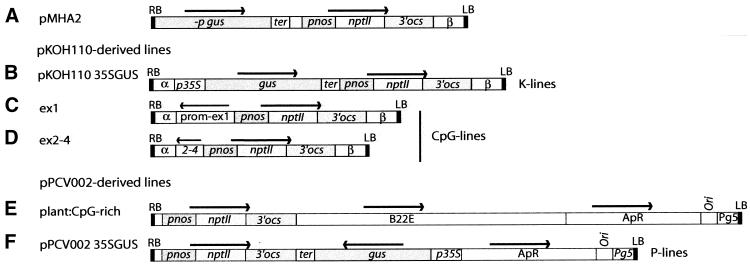
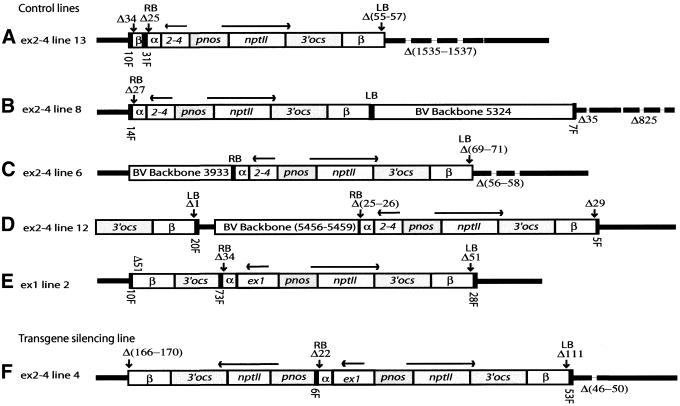
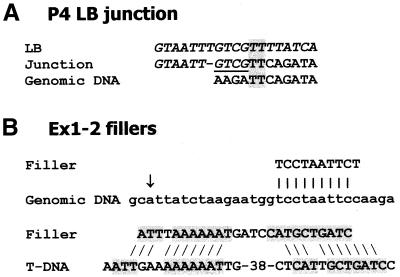
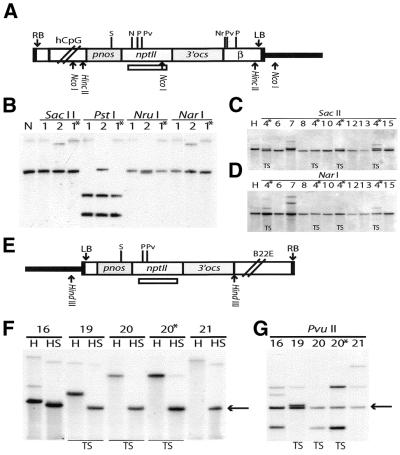
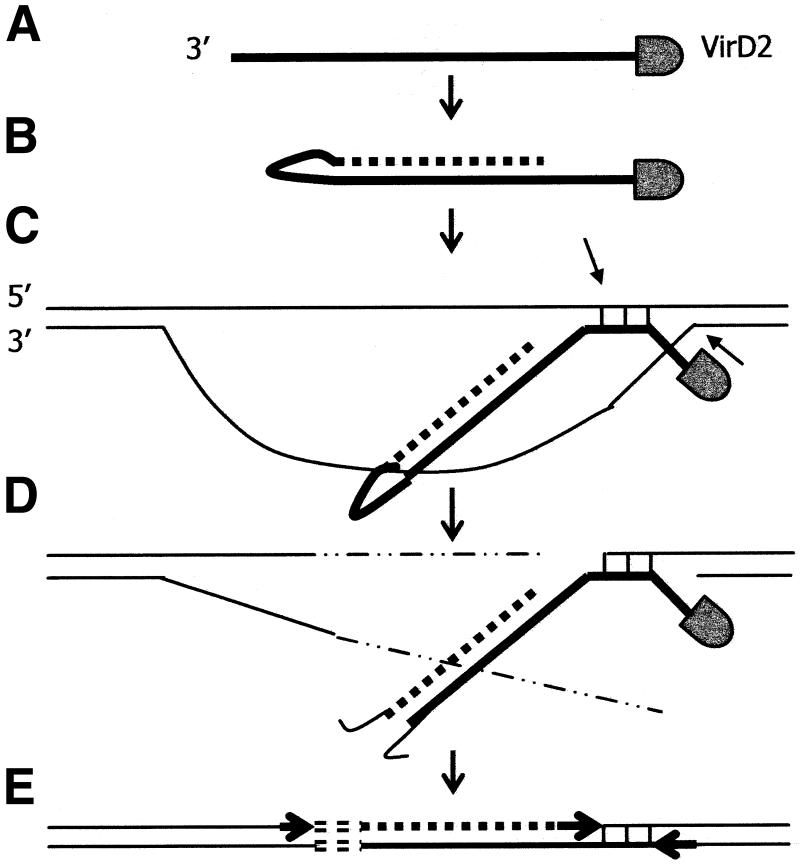
In genetically transformed plants, transgene silencing has been correlated with multiple and complex insertions of foreign DNA, e.g. T-DNA and vector backbone sequences. Occasionally, single-copy transgenes also suffer transgene silencing. We have compared integration patterns and T-DNA/plant DNA junctions in a collection of 37 single-copy T-DNA-transformed Arabidopsis lines, of which 13 displayed silencing. Vector sequences were found integrated in five lines, but only one of these displayed silencing. Truncated T-DNA copies, positioned in inverse orientation to an intact T-DNA copy, were discovered in three lines. The whole nptII gene with pnos promoter was present in the truncated copy of one such line in which heavy silencing has been observed. In the two other lines no silencing has been observed over five generations. Thus, vector sequences and short additional T-DNA sequences are not sufficient or necessary to induce transgene silencing. DNA methylation of selected restriction endonuclease sites could not be correlated with silencing. Our collection of T-DNA/plant DNA junctions has also been used to evaluate current models of T-DNA integration. Data for some of our lines are compatible with T-DNA integration in double-strand breaks, while for others initial invasion of plant DNA by the left or by the right T-DNA end seem important.
Figures

Similar articles
-
An inverted repeat transgene with a structure that cannot generate double-stranded RNA, suffers silencing independent of DNA methylation.Transgenic Res. 2006 Aug;15(4):489-500. doi: 10.1007/s11248-006-0019-y. Transgenic Res. 2006. PMID: 16906449
-
Local DNA features affect RNA-directed transcriptional gene silencing and DNA methylation.Plant J. 2008 Jan;53(1):1-10. doi: 10.1111/j.1365-313X.2007.03311.x. Epub 2007 Oct 27. Plant J. 2008. PMID: 17971044
-
High frequency of single-copy T-DNA transformants produced by floral dip in CRE-expressing Arabidopsis plants.Plant J. 2009 Aug;59(4):517-27. doi: 10.1111/j.1365-313X.2009.03889.x. Epub 2009 Apr 6. Plant J. 2009. PMID: 19392707
-
Position effects and epigenetic silencing of plant transgenes.Curr Opin Plant Biol. 1998 Apr;1(2):142-8. doi: 10.1016/s1369-5266(98)80016-2. Curr Opin Plant Biol. 1998. PMID: 10066569 Review.
-
RNA-based silencing strategies in plants.Curr Opin Genet Dev. 2001 Apr;11(2):221-7. doi: 10.1016/s0959-437x(00)00183-0. Curr Opin Genet Dev. 2001. PMID: 11250148 Review.
Cited by
-
Number and accuracy of T-DNA insertions in transgenic banana (Musa spp.) plants characterized by an improved anchored PCR technique.Transgenic Res. 2006 Apr;15(2):139-50. doi: 10.1007/s11248-005-2544-5. Transgenic Res. 2006. PMID: 16604456
-
Epigenetic interactions between Arabidopsis transgenes: characterization in light of transgene integration sites.Plant Mol Biol. 2003 May;52(1):217-31. doi: 10.1023/a:1023941123149. Plant Mol Biol. 2003. PMID: 12825701
-
A Cytotoxic and Anti-inflammatory Campesterol Derivative from Genetically Transformed Hairy Roots of Lopezia racemosa Cav. (Onagraceae).Molecules. 2017 Jan 12;22(1):118. doi: 10.3390/molecules22010118. Molecules. 2017. PMID: 28085103 Free PMC article.
-
High frequency Agrobacterium tumefaciens-mediated plant transformation induced by ammonium nitrate.Plant Cell Rep. 2009 May;28(5):737-57. doi: 10.1007/s00299-009-0676-4. Epub 2009 Feb 17. Plant Cell Rep. 2009. PMID: 19221758
-
An inverted repeat transgene with a structure that cannot generate double-stranded RNA, suffers silencing independent of DNA methylation.Transgenic Res. 2006 Aug;15(4):489-500. doi: 10.1007/s11248-006-0019-y. Transgenic Res. 2006. PMID: 16906449
References
-
- Gheysen G., Angenon,G. and Van Montagu,M. (1998) Agrobacterium-mediated plant transformation: a scientifically intriguing story with significant applications. In Lindsey,K. (ed.), Transgenic Plant Research. Harwood Academic Publishers, Amsterdam, The Netherlands, pp. 1–33.
-
- Zupan J., Muth,T.R., Draper,O. and Zambryski,P. (2000) The transfer of DNA from Agrobacterium tumefaciens into plants: a feast of fundamental insights. Plant J., 23, 11–28. - PubMed
