

Alpha/Beta interferon and gamma interferon synergize to inhibit the replication of herpes simplex virus type 1
- PMID: 12388715
- PMCID: PMC136787
- DOI: 10.1128/jvi.76.22.11541-11550.2002
Alpha/Beta interferon and gamma interferon synergize to inhibit the replication of herpes simplex virus type 1
Abstract
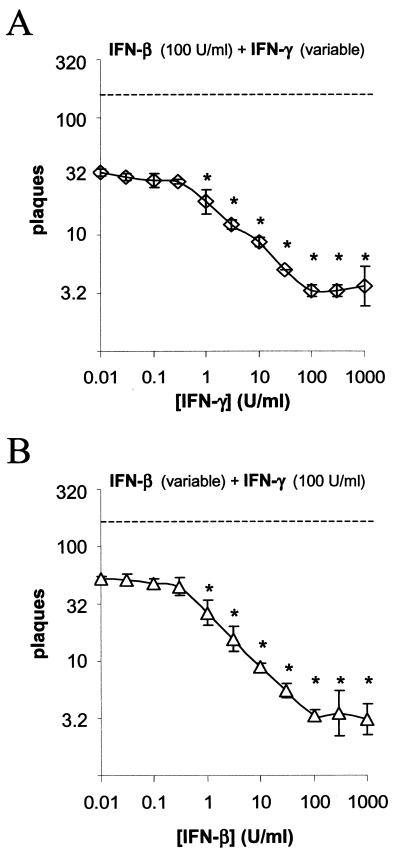
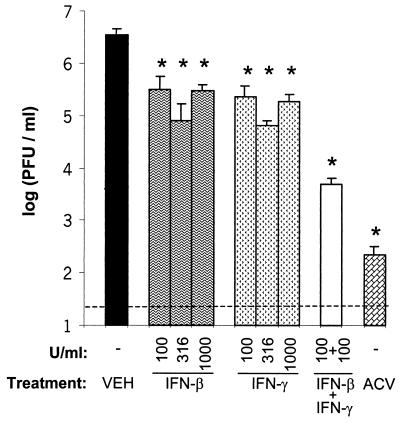
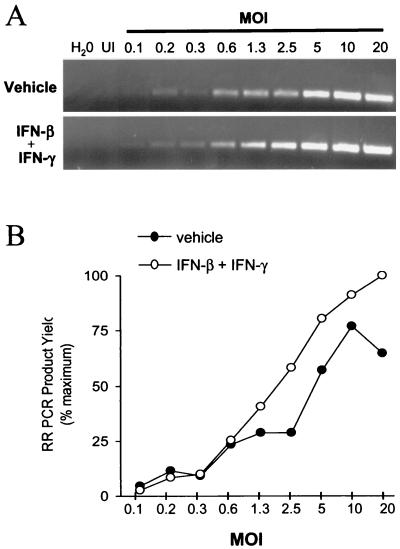
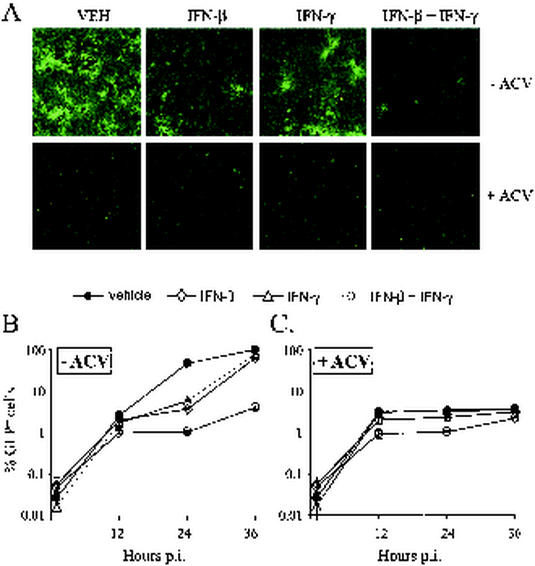
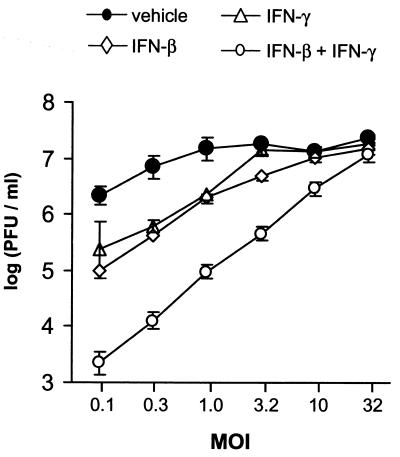
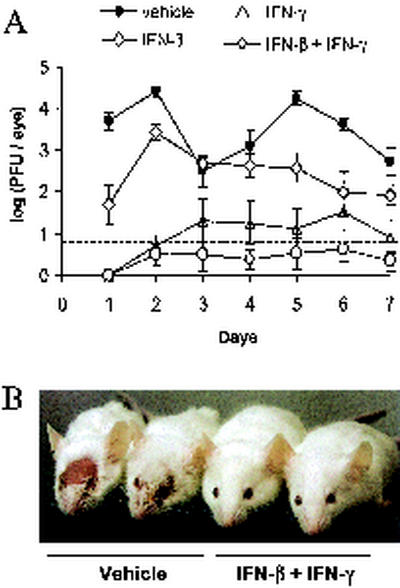
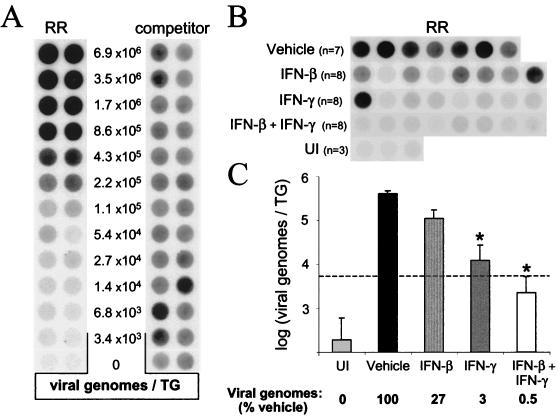
In vivo evidence suggests that T-cell-derived gamma interferon (IFN-gamma) can directly inhibit the replication of herpes simplex virus type 1 (HSV-1). However, IFN-gamma is a weak inhibitor of HSV-1 replication in vitro. We have found that IFN-gamma synergizes with the innate IFNs (IFN-alpha and -beta) to potently inhibit HSV-1 replication in vitro and in vivo. Treatment of Vero cells with either IFN-beta or IFN-gamma inhibits HSV-1 replication by <20-fold, whereas treatment with both IFN-beta and IFN-gamma inhibits HSV-1 replication by approximately 1,000-fold. Treatment with IFN-beta and IFN-gamma does not prevent HSV-1 entry into Vero cells, and the inhibitory effect can be overcome by increasing the multiplicity of HSV-1 infection. The capacity of IFN-beta and IFN-gamma to synergistically inhibit HSV-1 replication is not virus strain specific and has been observed in three different cell types. For two of the three virus strains tested, IFN-beta and IFN-gamma inhibit HSV-1 replication with a potency that approaches that achieved by a high dose of acyclovir. Pretreatment of mouse eyes with IFN-beta and IFN-gamma reduces HSV-1 replication to nearly undetectable levels, prevents the development of disease, and reduces the latent HSV-1 genome load per trigeminal ganglion by approximately 200-fold. Thus, simultaneous activation of IFN-alpha/beta receptors and IFN-gamma receptors appears to render cells highly resistant to the replication of HSV-1. Because IFN-alpha or IFN-beta is produced by most cells as an innate response to virus infection, the results imply that IFN-gamma secreted by T cells may provide a critical second signal that potently inhibits HSV-1 replication in vivo.
Figures
References
-
- Adler, H., J. L. Beland, N. C. Del-Pan, L. Kobzik, R. A. Sobel, and I. J. Rimm. 1999. In the absence of T cells, natural killer cells protect from mortality due to HSV-1 encephalitis. J. Neuroimmunol. 93:208-213. - PubMed
-
- Balish, M. J., M. E. Abrams, A. M. Pumfery, and C. R. Brandt. 1992. Enhanced inhibition of herpes simplex virus type 1 growth in human corneal fibroblasts by combinations of interferon-alpha and -gamma. J. Infect. Dis. 166:1401-1403. - PubMed
-
- Buddingh, G. J., D. I. Schrum, J. C. Lanier, and D. J. Guidry. 1953. Studies of the natural history of herpes simplex virus infections. Pediatrics 11:593-605. - PubMed
-
- Cantell, K. 1995. Development of antiviral therapy with alpha interferons: promises, false hopes, and accomplishments. Ann. Med. 27:23-28. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
