

RNA polymerase III in Cajal bodies and lampbrush chromosomes of the Xenopus oocyte nucleus
- PMID: 12388750
- PMCID: PMC129959
- DOI: 10.1091/mbc.e02-05-0281
RNA polymerase III in Cajal bodies and lampbrush chromosomes of the Xenopus oocyte nucleus
Abstract
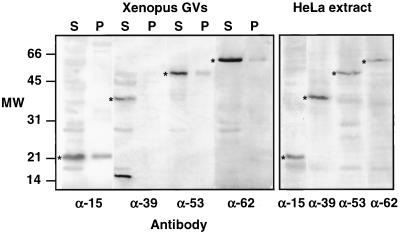
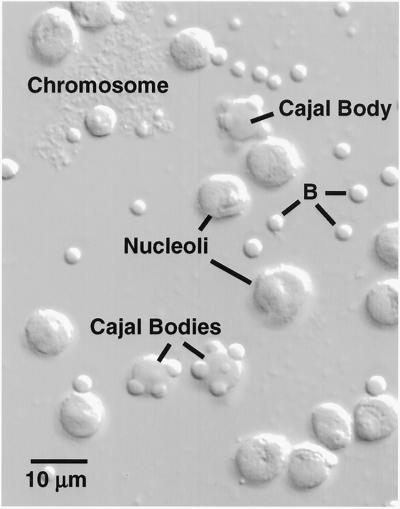
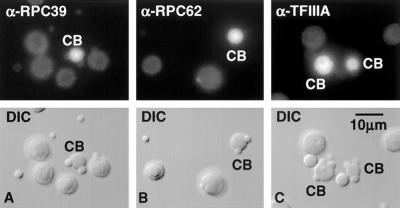
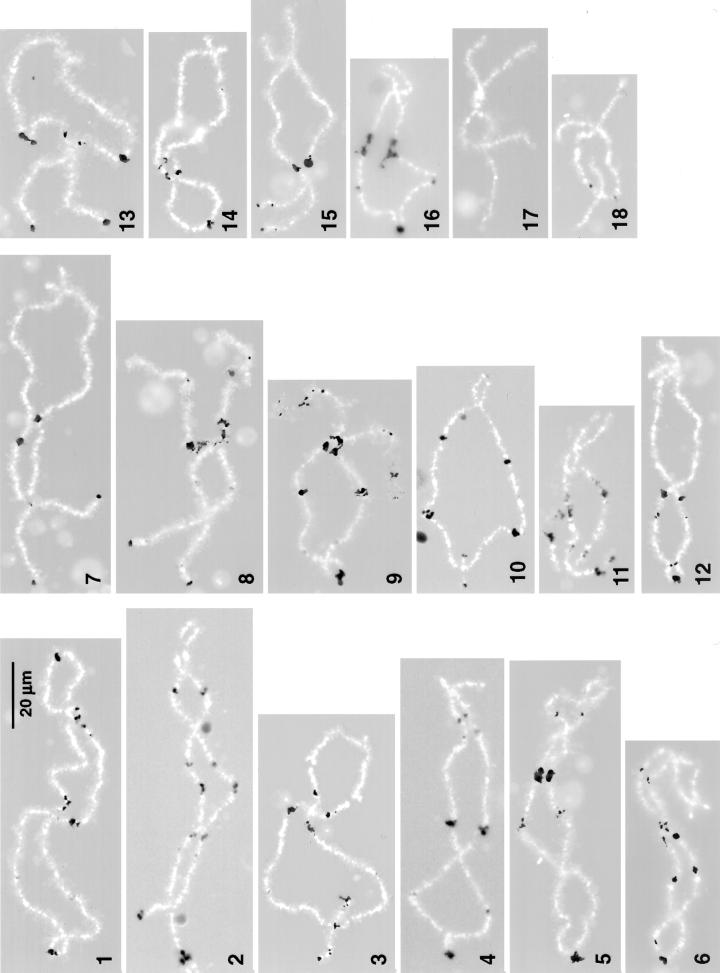
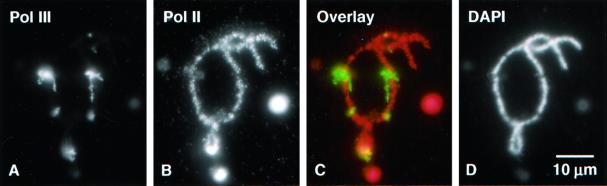
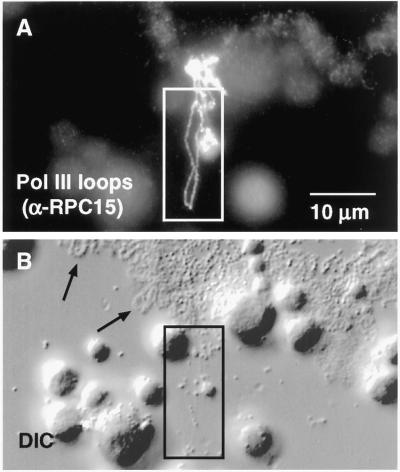
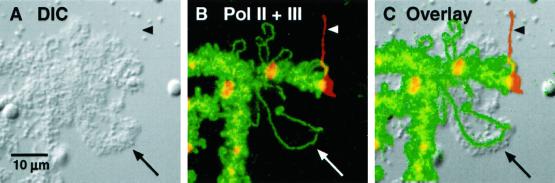
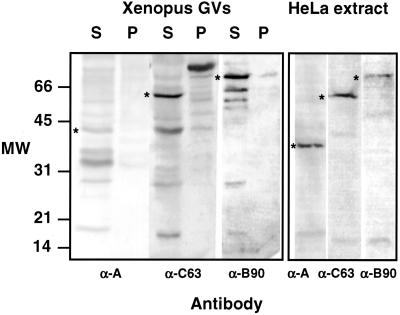
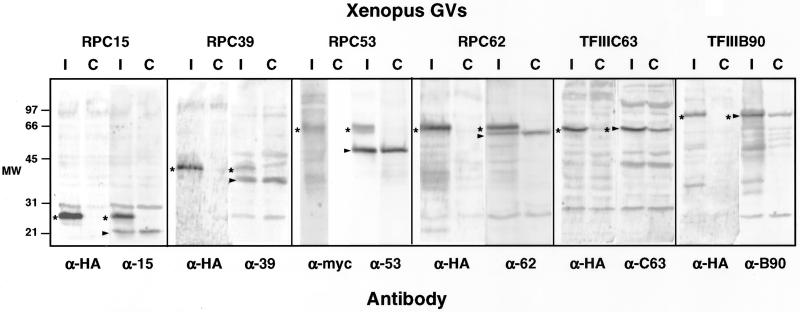
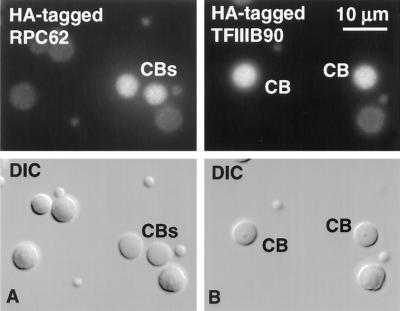
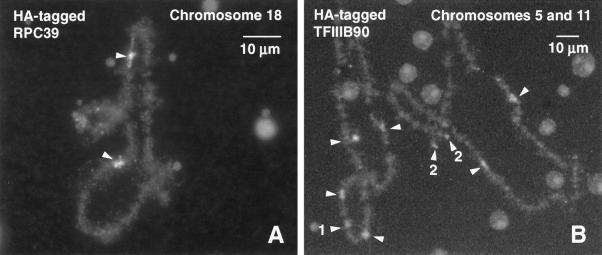
We used immunofluorescence to study the distribution and targeting of RNA polymerase (pol) III subunits and pol III transcription factors in the Xenopus laevis oocyte nucleus. Antibodies against several of these proteins stained Cajal bodies and approximately 90 specific sites on the lampbrush chromosomes. Some of the chromosomal sites had been identified previously by in situ hybridization as the genes for 5S rRNA. The remaining sites presumably encode tRNAs and other pol III transcripts. Pol III sites were often resolvable as loops similar to the much more abundant pol II loops, but without a matrix detectable by phase contrast or differential interference contrast. This morphology is consistent with the transcription of short repeated sequences. Hemagglutinin-tagged transcripts encoding core subunits and transcription factors were injected into the oocyte cytoplasm, and the distribution of newly translated proteins inside the nucleus was monitored by immunostaining. Cajal bodies were preferentially targeted by these proteins, and in some cases the chromosomal sites were also weakly stained. The existence of pol III subunits and pol III transcription factors in Cajal bodies and their targeting to these organelles are consistent with a model of Cajal bodies as sites for preassembly of the nuclear transcription machinery.
Figures
References
-
- Acker J, Wintzerith M, Vigneron M, Kedinger C. A 14.4 kDa acidic subunit of human RNA polymerase II with a putative leucine-zipper. DNA Seq. 1994;4:329–331. - PubMed
-
- Angelier N, Lacroix J-C. Complexes de transcription d'origines nucléolaire et chromosomique d'ovocytes de Pleurodeles waltlii et P. poireti (Amphibiens, Urodèles) Chromosoma. 1975;51:323–335. - PubMed
-
- Brown DD, Sugimoto K. 5S DNAs of Xenopus laevis and Xenopus mulleri: evolution of a gene family. J Mol Biol. 1973;78:397–415. - PubMed
-
- Bucci S, Nardi I, Mancino G, Fiume L. Incorporation of tritiated uridine in nuclei of Triturus oocytes treated with α-amanitin. Exp Cell Res. 1971;69:462–465. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
